Sensibilité du modèle aux priors informatifs
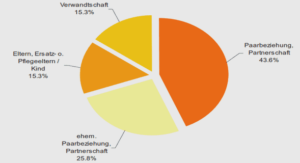
Le modèle a été utilisé avec succès au Danemark, en Grande-Bretagne et aux Pays-Bas. Cependant, les résultats obtenus pour ces pays sont apparus comme divergents quant aux sources principales, hors cas liés à un voyage. De plus, pour la Pologne et l’Allemagne, la convergence du modèle n’a pu être atteinte. La raison invoquée repose sur le manque de qualité et de représentativité des données, ce qui semble peu réaliste pour les données allemandes. Le modèle proposé par les Danois est un modèle surparamétré et nécessite donc l’introduction d’information a priori « informative » sur certains paramètres. Cette information informative peut porter sur le paramètre source-dépendant (qui mesure la différence entre les sources quant à leur capacité à véhiculer les salmonelles), et/ou sur le paramètre type-dépendant (qui mesure la différence entre les types quant à leur capacité à causer des infections). L’objectif des travaux présentés ci-après était d’évaluer l’impact de l’information informative introduite dans le modèle. Pour ce faire, les analyses ont été conduites à partir du jeu de données français, pour lequel seules 4 sources potentielles ont été considérées (poules pondeuses, poulets de chair, dindes et porcs). Différentes variantes du modèle ont été considérées : un modèle simple déterministe, qui sert de base de référence, le modèle originel assorti de quatre types d’information différents, ainsi que l’adaptation du modèle proposée par Mullner et al (2009)1 . Les quatre types d’information considérés correspondent à deux variantes de la paramétrisation utilisée dans la publication originelle (Hald, Vose et al. 2004) (le paramètre type-dépendant est fixé à une valeur arbitraire pour un type de référence), et deux variantes que nous proposons (les paramètres type-dépendants correspondants aux types spécifiques sont fixés soit à une valeur arbitraire soit à une valeur calculée à partir des données). Les résultats obtenus pour chacune de ces 6 variantes sont comparés en termes d’adéquation du modèle, de résultats d’attribution par source et de distribution a posteriori des paramètres source et type dépendants. Le projet d’article suivant est prévu pour soumission à Foodborne Pathogens and Disease, après relecture par un traducteur.
Data
A total of 9 076 human cases were included in the dataset, as well as the prevalence results for 519 layer farms, 371 broiler farms, 331 turkey farms, 1 166 pig carcasses and 334 cattle carcasses (Figure 1). However, because of the very low prevalence of Salmonella in cattle (only 2.4%), inducing an important asymmetric density of the marginal posterior distribution with a small number of attributed cases (around 50), cattle was excluded as a source for this comparative study. The types are heterogeneously distributed among the sources according to Fisher exact tests. Serotypes considered were those who were the most frequent among human cases (> 30 cases) and in sources (> 15% of the strains within a source) and those corresponding to specific types. The serotypes which didn’t fulfil those criteria were included in “others” category. For Enteritidis and Typhimurium, 9 subtypes were defined each. But only 5 subtypes for Typhimurium and 3 for Enteritidis were included in the model. Those subtypes are the ones observed simultaneously in the human cases and at least in one animal source. The other subtypes were grouped in categories “other Typhimurium” and “other Enteritidis”. Human cases belonging to the three “other” categories could not be considered for the attribution because, depending on the source, those categories have different types 155 compositions, which makes it impossible to distribute the relative cases according to prevalences that are not comparable. Thus, as these cases cannot be attributed to a source, 5 938 cases spread between 28 serotypes, 5 Typhimurium subtypes and 3 Enteritidis subtypes were considered for attribution and referred to as attributable cases. Our studied sample contains two reference types including 8 subtypes (2 of which are specific), 12 specific types not included in the reference types and 14 other serotypes. Excluding Enteritidis and Typhimurium subtypes, a total of 5 types are specific to pigs, 2 to layers and 2 to broilers and 2 to turkeys (Table 2). Finally, on the basis of the updated results of the INCA study, national consumption for 2005 was 82 301 tons for layers (eggs), 84 842 tons for broilers, 18 967 tons for turkeys and 161 971 tons for pigs.
Attribution
Results presented correspond to runs of 100 000 iterations of the Gibbs sampler with a thin of 25. Convergence diagnostics were satisfactory (Cowles and Carlin 1996; Brooks and Gelman 1998; Brooks and Roberts 1998b). From these runs, parameters estimates (posterior means, posterior variances and posterior 95% credibility intervals) were computed from the last 50 000 iterations. We first performed comparisons corresponding to adequacy diagnosis. It concerned the number of cases per type and the total number of attributed cases, which were compared to the observed values. We then analyzed the expected numbers of cases per source with the simple model’s results as reference. Finally, for models giving adequate results, the type and source dependant parameters were considered.
Adequacy of the model
Predicted and observed numbers of cases per type are in good agreement for Specific-Types 2 and for Mullner models, unlike both Reference-Type configurations and Specific-Types 1 (figure 2). Indeed, for both Reference-Type configurations, 5 subtypes are not well predicted. Regarding Typhimurium, subtypes 1, 2, 3 and 4 are concerned. The expected number of cases for Typhimurium globally is 113 for Reference-Type and 1 for ReferenceType 2, where 1 807 Typhimurium cases are observed. The discrepancy is less spectacular but still significant for Enteritidis, with 2 concerned subtypes, SE multiS and SE1. For Reference-Type 1, 1 796 and 869 cases are expected respectively for SE-multiS and SE1 vs 2 092 and 615 observed cases. For Reference-Type 2, 315 cases are expected for SE1 vs 615 observed. These weak fits are not related to the cut off of the marginal posteriors distributions of the q’s, except for SE1 in the Reference-Type 2 configuration. For Specific-Types 1 configuration, almost all specific types are not well predicted (Stourbridge, Bovismorbificans, Oranienburg, Heidelberg, S 48:z4,z23:-, Havana, Ohio, Goldcoast, Bareilly, Aijobo). Though, in this last case, the convergence is good and the mismatch concerns only serotypes associated to a small number of cases (5% of attributable cases overall). As a consequence, the total expected number of cases with Specific-Types 2 (n= 5 745) and Mullner (n = 6 000) models are in accordance with the 5 938 observed attributable domestic sporadic cases. This is also true for Specific-Types 1 (n = 5 745) despite the discrepancy for some of the specific types. Finally, for the Reference-Type configurations, a decrease of around 30% in attributed cases is observed (4 122 expected cases for Reference-Type 1 and 4 079, for Reference-Type 2).