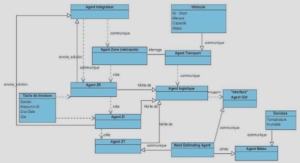
Productivité forestière et modèles conceptuels
Il existe plusieurs modèles conceptuels qui mettent en relation la productivité forestière et les facteurs influençant cette dernière. À titre d’exemple, Bonan (1989) a mis en place un schéma conceptuel (Figure 1.1) dans un modèle de simulation de la dynamique de la forêt boréale qui regroupe les facteurs intrinsèques et extrinsèques pouvant influencer la régénération, la croissance et la mortalité des arbres dans les forêts de conifères, de feuillus et mixtes dans plusieurs sous-régions bioclimatiques de la zone boréale nord-américaine. Ces facteurs comprenaient le climat, le rayonnement solaire, l’humidité du sol, la température du sol, la couche organique du sol forestier, la disponibilité des nutriments, les incendies de forêt et les épidémies d’insectes. Également, dans les schémas conceptuels d’Austin et Smith (1990) et Barnes et al. (1997), les facteurs environnementaux (intrinsèques et extrinsèques) ont un effet direct sur les processus physiologiques des arbres et sur leur réponse en regard de la productivité (Figure 1.2).
Dans la forêt boréale aménagée, il existe d’autres facteurs, dits « facteurs d’aménagement » qui peuvent interagir avec d’autres facteurs intrinsèques (p. ex. sol forestier) et influencer significativement la productivité forestière à l’échelle du site. Ces facteurs sont la fertilisation, la sélection génétique des essences, le contrôle de la végétation concurrente, le contrôle de la densité de reboisement, la taille et l’état des plants (Figure 1.3) (Richardson et al., 1999). À titre d’exemple, en forêt boréale russe, on a fixé une certaine densité de plants de pins pour maintenir une productivité optimale dans les zones à régénérer. Le nombre recommandé de plants est de 3 000 à 4 000 par hectare (Karvinen et al., 2011). Par ailleurs, en Alaska, les plus faibles taux de survie de l’épinette blanche (25% à 40%, faible productivité) étaient liés à la présence de végétation concurrente, telle que la calamagrostide du Canada (Calamagrostis canadensis (Michx.) Beauv.) et l’épilobe à feuilles étroites (Chamaenerion angustifolium (L.) Scopoli subsp. angustifolium) (Graham et Wurtz, 2003). Des études réalisées dans des sites reboisés en forêt boréale canadienne ont montré que les plants de taille importante avaient des taux de survie et de croissance plus élevés que les petites plantules et étaient susceptibles de maximiser la productivité forestière du futur peuplement (Hamel et al., 2004; Sutherland et Day, 1988; Thomson et McMinn, 1989). Généralement, les caractéristiques du sol (fertilité, texture, structure, humidité) sont parmi les facteurs les plus influents, car elles peuvent influencer significativement la productivité forestière du site. Donc, toute pratique d’aménagement pouvant détériorer le continuum sol-plante aura des conséquences néfastes (p. ex. érosion, lessivage des nutriments, compactage du sol, envahissement du site par la végétation concurrente) sur la productivité future du site (Austin et Smith, 1990; Bonan, 1989; Richardson et al., 1999).
Forêts paludifiées de l’Est canadien
La forêt boréale de l’Est canadien présente un fort potentiel de production ligneuse, notamment dans le domaine bioclimatique de la pessière noire à mousses. Toutefois, cette région est caractérisée par une faible productivité dans les sites paludifiés. La paludification (entourbement) est un phénomène naturel où l’épaisseur de la couche organique augmente graduellement jusqu’à excéder, voire largement dépasser 30 cm menant graduellement à la formation d’une forêt tourbeuse (Bernier et al., 2008; Payette et Rochefort, 2001). Ce milieu est caractérisé par une saturation en eau du sol, conduisant à un milieu anaérobique froid, qui diminue l’activité microbiologique (Comont, 2006). Ces conditions limitent la minéralisation des éléments nutritifs et leur absorption par les plantes (Gower et al., 1996; Murty et al., 1996; Prescott et al., 2000) causant une diminution de la croissance et une hausse de la mortalité des arbres en place, ainsi qu’une faible régénération (Simard et al., 2009, 2007). Selon Fenton et al. (2005, 2010) et Greene et al. (2006), le processus de paludification est lié à une multitude de facteurs qui peuvent le contrôler ou l’accentuer. La sévérité des feux de forêt et leurs fréquences, le type de peuplement, la topographie et le niveau de la pente sont parmi ces facteurs. Par exemple, les feux sévères et les pentes inclinées (bon drainage) permettent de freiner le processus de la paludification (Simard et al., 2007). Selon Payette et Rochefort (2001), il existe deux principaux types de processus de paludification, la paludification édaphique et la paludification successionelle. La paludification édaphique, également appelée paludification des dépressions humides, opère où la topographie joue un rôle déterminant dans le maintien de la nappe phréatique proche de la surface du sol. La paludification successionelle, également appelée paludification des sites bien drainés, opère où la succession forestière est le principal moteur du processus (Simard et al., 2007). Sur le terrain, ces deux processus sont confondus mais leurs différences sont importantes pour l’aménagement forestier (Laamrani et al., 2004a, b). La paludification successionelle est un phénomène potentiellement réversible, tandis que la paludification édaphique est fortement liée aux propriétés intrinsèques du site dont la modification nécessiterait des ressources considérables (p. ex. drainage forestier) (Simard et al., 2009).
La paludification est très répandue dans la pessière à mousses de l’Est canadien, notamment dans les pessières à mousses de l’Ouest québécois en raison du relief plat, du climat froid et à la présence de sols argileux à drainage lent (Gauthier et Vaillancourt, 2008; Mansuy et al., 2018). Ces conditions diminuent la capacité de drainage (conditions hydromorphes et anaérobiques) et l’activité microbienne à cause des conditions d’anaérobies du sol. À leur tour, ces conditions diminuent le taux de décomposition de la matière organique accumulée sur le sol argileux (Fenton et al., 2005). La paludification crée des conditions défavorables pour la croissance de la régénération de l’épinette noire due à la mauvaise qualité des substrats organiques (microsites) surfaciques dominés par un tapis de sphaignes, souvent gorgées d’eau et pauvres en éléments nutritifs (Lavoie et al., 2007a; Fenton et al., 2005). La productivité ligneuse des pessières noires peut diminuer de 50 à 80 % sur plusieurs centaines d’années; cette perte de productivité s’observe particulièrement entre 100 et 200 ans après feu (Drobyshev et al., 2010; Simard et al., 2009, 2007). Durant cette période, l’épaisseur de la couche organique passe graduellement de 20 à 40 cm. La zone d’enracinement de l’épinette noire, laquelle est superficielle, migre alors du sol minéral à la couche organique saturée et pauvre en éléments nutritifs (Lavoie et al., 2007a, 2007b; Simard et al., 2007).
Dans les pessières à mousses de l’Est canadien, les perturbations des pratiques sylvicoles actuelles ne permettent pas de limiter le processus de paludification. Parmi ces pratiques sylvicoles, on note la coupe avec protection de la régénération et des sols (CPRS), fréquemment utilisée au Québec, qui conserve la couche organique et le couvert de sphaignes entre les sentiers de débardage (Bernier et al., 2008; Lafleur et al., 2010a, b). Les coupes hivernales sont particulièrement problématiques dans les sites paludifiés, car la couche de neige ne permet pas de perturber (et de mélanger) le sol sous-jacent (Lafleur et al., 2010a). De plus, durant la saison estivale, ces sites récoltés sont difficilement accessibles pour la remise en production via le reboisement, à cause des conditions hydromorphes du sol (orniérage) qui limitent la circulation de la machinerie sylvicole (p. ex. enfouissement des roues dans les sols très humides). Néanmoins, dans les sites paludifiés, les sylviculteurs doivent déployer des efforts importants (p. ex. préparation mécanique du sol (PMS)) pour assurer la remise en production de ces sites, notamment en augmentant la disponibilité des microsites propices à la croissance (Bergeron et al., 2007; Lavoie et al., 2005).
Objectifs de la thèse
L’objectif général de cette thèse est de déterminer les effets de certains facteurs édaphiques (teneur en eau du sol, disponibilité des éléments nutritifs du sol nécessaires à la croissance : N, P, K, Ca, Mg, épaisseur de la couche organique) et certains facteurs environnementaux et topographiques (disponibilité et qualité des microsites, compétition interspécifique, pente, orientation, position d’établissement du plant) sur la survie et la croissance (hauteur et diamètre) des plants de reboisement installés sur des sites ayant subi une altération des horizons organiques et minéraux par différentes intensités de perturbations créées par plusieurs traitements sylvicoles. Pour ce faire, l’analyse se réalisera sur trois échelles, notamment à l’échelle du microsite, à l’échelle du peuplement et à l’échelle régionale. Ces travaux permettront, à terme, de mieux comprendre les conditions d’établissement de la régénération artificielle en forêt boréale de l’Est canadien, notamment en forêts paludifiées; et les interactions entre la croissance des plants et leur environnement (à l’échelle du microsite, du peuplement et de la région) suite à leur mise en terre, et ce, afin d’assurer le maintien de la productivité des sites boréaux et paludifiés récoltés. Le chapitre II de la thèse a pour objectif d’évaluer la disponibilité et la qualité des microsites (incluant le type de substrats) influençant l’établissement et la croissance des plants d’épinette noire mis en terre sur des sites paludifés suite aux perturbations générées par trois traitements sylvicoles (coupe totale (CPRS), PMS par le scarificateur mécanique T26, PMS par la herse forestière). Dans le chapitre III, nous avons déterminé l’effet des substrats minéraux, organiques et organo-minéraux sur la croissance des plants d’épinette noire en milieu contrôlé. Dans le chapitre IV, nous avons identifié les facteurs climatiques (échelle régionale), édaphiques (échelle du peuplement) et locaux (échelle du microsites) influençant la survie et la croissance initiale des plants d’épinette noire Picea mariana (Mill.) B.S.P et de pin gris (Pinus banksiana Lamb.). Dans le chapitre V, nous récapitulons la contribution de la thèse dans l’avancement de connaissances et proposons des pistes d’aménagement forestier et de sylviculture qui permettent de garantir de meilleures conditions d’établissement des plants dans les sites boréaux aménagés et une remise en production efficace des sites paludifiés de l’Est canadien.
AVANT-PROPOS |