Étude de la circulation de la fièvre Q, de l’ADN de Coxiella burnetii et Borrelia spp. chez l’Homme
Présentation de la bactérie Coxiella burnetii
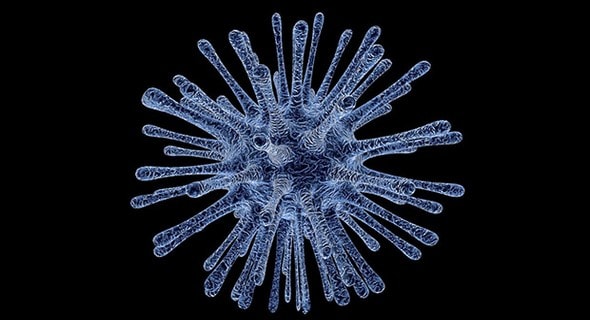
Coxiella burnetii est une coccobacille Gram négative (0,2 à 2 μm) (Fournier et al., 1998 ; Van Leuken et al., 2016). Elle partage de nombreuses propriétés physiologiques avec les Rickettsies (Boarbi et al., 2015). C’est un pathogène intracellulaire qui se réplique dans les cellules eucaryotes (Van den Brom et al., 2015). Sa vacuole de réplication acquiert progressivement les caractéristiques d’un phagolysosome (pH acide, hydrolysats acides et peptides cationiques). La bactérie participe activement à la genèse de cette vacuole intracellulaire et dispose de plusieurs stratégies d’adaptation à cet environnement acide (Eldin et al., 2017). Figure 1 : Bactérie Coxiella burnetii, agent de la fièvre Q (Photo D. Raoult)
Description et position systématique de C. burnetii
Des études phylogénétiques récentes, basées sur la séquence du gène codant l’ARN ribosomal 16S, ont montré que le genre Coxiella devrait être exclu de l’ordre des Rickettsiales car il appartiendrait à l’ordre des Legionellales. Figure 2 : Classification de C. burnetii, D’après Seshadri et al., 2003 ; Arricau-Bouvery & Rodolakis, 2005 Phylum Classe Ordre Famille Proteobacteria Gammaproteobacteria Legionellales Domaine Bacteria Coxiellaceae
Des travaux de Rousset et collaborateurs (2007) ont indiqué que le genre Coxiella ne renferme qu’une seule espèce C. burnetii. Mais, une bactérie isolée d’écrevisses australiennes (Cherax quadricarinatus) nommée Coxiella cheraxi proche génétiquement de C. burnetii a été décrite (Tan & Owens, 2000).
Caractères biologiques et variations génétiques de C. burnetii
La caractérisation de C. burnetii a été réalisée par la mise au point de sa culture axénique (Eldin et al., 2017). C. burnetii est également cultivable sur œufs embryonnés, lignées macrophagiques ou cellules Vero (Boarbi et al., 2015). L’étude des plasmides a permis de déterminer 4 à 6 génotypes qui seraient associés à des lipopolysaccharides de surface (LPS) différents voire des variations de leur pouvoir pathogène. Les variations génétiques semblent être en relation avec l’origine géographique des souches et non avec leur pouvoir pathogène. Les variations de phases antigéniques sont liées à des modifications du LPS (Rousset et al., 2007). La phase I virulente qui possède un LPS complet est la forme naturelle de la bactérie et elle ne doit être manipulée que dans un laboratoire de sécurité de niveau 3. La phase II possède un LPS incomplet causé par une délétion de gènes dans le chromosome entrainant une perte de certaines protéines de la membrane externe (Rousset et al., 2007) et son pouvoir infectieux est faible.
Cycle intracellulaire et différenciation morphologique de C. burnetii
Cycle intracellulaire et variation de phase
Le cycle intracellulaire de C. burnetii est complexe. Les bactéries en phase I pénètrent dans les cellules cibles en se fixant sur des récepteurs apparentés aux intégrines (Boarbi et al., 2015). Après la phagocytose, les bactéries sont présentes dans les phagosomes qui fusionnent avec les lysosomes pour former des phagolysosomes. Ces derniers fusionnent entre eux pour former une vacuole unique dont le pH est acide (Coleman et al., 2004) et varie entre 4,7-5,2. Cette acidité nécessaire au métabolisme et à la multiplication du microorganisme entrave l’efficacité de certains antibiotiques lors du traitement (Malosse, 2008). Les bactéries en phase II présentent un comportement très différent car elles se fixent sur les récepteurs du fragment iC3b du système complémentaire (CR3) (Raoult et al.,2005 ; Arricau-Bouvery & Rodolakis 2005), ou elles sont rapidement détruites par les phagolysosomes. Etude de la circulation de la fièvre Q, de l’ADN de Coxiella burnetii et Borrelia spp. Chez l’Homme, les tiques et dans le milieu domestique en zones rurales au Sénégal Master Biologie Animale FST-UCAD, 2018 5 Ces différences de comportement entre C. burnetii en phase I et II permettent d’expliquer que seules les bactéries en phase I soient infectieuses.
Différenciation cellulaire
La multiplication cellulaire de C. burnetii aboutit à la formation de deux formes différentes par leur morphologie, leurs caractères biochimiques et leur métabolisme (ArricauBouvery & Rodolakis, 2005 ; Eldin et al., 2013) : les variantes de petite taille ou Small Cell Variant (SCV) et les variantes de grande taille ou Large Cell Variant (LCV). Les SCV mesurent 0,2 à 0,5 µm (Coleman et al., 2004) et sont denses en microscopie électronique (McCaul et Williams, 1981 ; Eldin et al., 2017). Ce sont les formes capables de résister dans l’environnement et sont métaboliquement peu actifs (Heinzen et al.,1999). Sous cette forme, C. burnetii peut être présente pendant plusieurs années dans la poussière (Tissot-Dupont et al., 2004). Métaboliquement très actifs, les formes LCV représentées par de grosses cellules de forme allongée, mesurant 0,7 à 2 µm sont polymorphes, peu denses et exclusivement intracellulaires. Elles sont infectieuses mais très fragiles dans le milieu extracellulaire (McCaul et Williams, 1981), ils sont activés par l’acidité des phagolysosomes et elles sont responsables de la dissémination de C. burnetii dans un organisme infecté. Au cours de leur multiplication, les LCV semblent capables de présenter un phénomène proche de celui de la sporulation qui les divisera en deux compartiments de taille inégale dont chacun renferme un matériel nucléaire complet (Malosse, 2008). Le petit compartiment cellulaire donnerait naissance à une pseudo-spore dont le développement conduirait à une variante de petite taille (McCaul & Williams, 1981).
Pouvoir pathogène de C. burnetii
Coxiella burnetii est une bactérie possédant un pouvoir infectieux élevé (Van Leuken et al., 2016). L’inoculation de deux bactéries en phase I à des souris, cobayes, œufs embryonnés ou cultures cellulaires assure l’infection de 55% de ces organismes, soit une ID50 de 0,5 à 2 (Van Leuken et al., 2016). La forte virulence de C. burnetii, la possibilité de son aérosolisation et sa stabilité environnementale ont conduit le CDC (Center for Disease Control and Prevention) des États-Unis à classer cette bactérie comme un agent de menace biologique de catégorie B (Raoult et al., 2005 ; Eldin et al., 2017). Une attaque de bioterrorisme avec ce pathogène pourrait entraîner des conséquences à long terme en raison de la possibilité de provoquer une infection persistante au sein de la population (Kersh et al., 2010).
Modes de transmission de C. burnetii
Transmission par voie aérienne
Chez l’homme, la transmission par voie respiratoire est la plus courante (Angelakis & Raoult, 2010). Dans les conditions expérimentales, l’inhalation d’une seule bactérie peut provoquer l’infection et la maladie chez l’homme (Pommier de Santi, 2016). Les mises-bas génèrent un risque maximal de diffusion de la bactérie dans l’environnement (Tissot-Dupont et al., 2004). Une fois excrétées, les spores de C. burnetii peuvent être transportées par le vent sous forme d’aérosols sur une distance de plus de 40 km et peuvent survivre pendant plusieurs années dans la poussière (Tissot-Dupont et al., 2004).
Autres modes de transmission
Les tiques peuvent transmettre C. burnetii à l’homme pendant le repas de sang, même si la transmission par piqûre de tiques est rare (Angelakis & Raoult, 2010). Elles peuvent excréter la bactérie par leurs excréments, la salive et le liquide coxal (Eldin et al., 2017). La transmission peut également se produire par contact direct ou indirect avec des animaux infectés ou leurs excréments séchés, avec de l’engrais et le linge des personnes exposées (Eldin et al., 2017). La transmission sexuelle a été démontrée chez des animaux de laboratoire. Il a été possible, à partir d’une semence de taureau naturellement infecté, d’isoler la bactérie par inoculation à la souris (Angelakis & Raoult, 2010). Etude de la circulation de la fièvre Q, de l’ADN de Coxiella burnetii et Borrelia spp. Chez l’Homme, les tiques et dans le milieu domestique en zones rurales au Sénégal Master Biologie Animale FST-UCAD, 2018 7 La voie orale pourrait être un mode important puisque les animaux peuvent se contaminer par léchage ou l’ingestion des produits de la parturition (Boarbi et al., 2015). La contamination par la voie alimentaire reste encore controversée car la dose infectieuse requise est assez méconnue. Cependant, cette voie suscite certaines précautions pour protéger les consommateurs (Rodolakis et al., 2004).
Le réservoir animal de C. burnetii
Les ruminants domestiques (ovins, bovins, caprins) sont les principaux réservoirs responsables des épidémies humaines liées à C. burnetii (Pommier de Santi et al., 2016). Cependant, l’identification des animaux infectés est difficile, car les animaux laitiers peuvent excréter la bactérie dans l’environnement sans être symptomatiques. Les principales manifestations cliniques observées chez les ruminants infectés sont les troubles de la reproduction tels que l’infertilité, la mortinatalité, l’avortement et l’endométrite (Van den Brom et al., 2015). Les femelles infectées excrètent une énorme quantité de bactéries dans les produits de naissance, l’urine, les excréments et le lait (Duron et al., 2015 ; Van den Brom et al., 2015). C. burnetii peut infecter d’autres mammifères domestiques (chevaux, chameaux, chiens, chats, lapins), sauvages (cervidés, ours, renards, marsupiaux, rongeurs), marins mais également les oiseaux (Boarbi et al., 2015)
INTRODUCTION |