Les porphyrines dicarboxyliques et la deutéroporphyrine
Structure
La deutéroporphyrine (DP, masse molaire 512 Da) est constituée du noyau porphyrique sur lequel sont greffés, du même côté et en position β, deux groupes carboxyliques (figure 1.1). Selon la nomenclature internationale, il s’agit en fait du « 2,7,12,18 tetramethylporphyrin –13,17 dipropionic acid ». Cette molécule présente un fort caractère amphiphile, avec une répartition asymétrique des zones hydrophobes et polaires. Elle peut être considérée comme l’archétype des porphyrines dicarboxyliques.
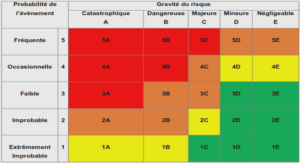
La deutéroporphyrine peut présenter divers états d’ionisation. En effet, le pH joue sur l’état de protonation des cycles pyrroles et des groupes périphériques. Ainsi, à pH acide, un ou deux protons peuvent se fixer aux azotes centraux et donner respectivement les formes monocationiques et dicationiques. A contrario, la déprotonation des groupes carboxyliques conduit aux formes monoanionique et dianionique (figure 1.2). Les pK des différents groupes ont été déterminés ([13] et tableau 1.1). Cependant, la précision des valeurs est assez faible, surtout concernant les groupes carboxyliques pour lesquels les phénomènes observés sont vraisemblablement dus en partie à la formation d’agrégats. Une partie de ce travail de thèse a donc consisté en la détermination précise de ces pK, notamment par des mesures dynamiques au stopped-flow. Notons enfin que les azotes pyrroliques ne peuvent se déprotoner qu’à des pH extrêmement forts (pK>16), incompatibles avec les systèmes biologiques que nous étudions.
Enfin, les solutions aqueuses de porphyrines sont relativement instables. En outre, des phénomènes d’autoassociation des molécules sont responsables de la formation de dimères ou éventuellement d’agrégats à des concentrations en porphyrine plus importantes. Par exemple, la constante de dimérisation de la deutéroporphyrine à 25°C dans du tampon phosphate à pH 7,2 est de 2,3×106 M-1 [99]. Cette valeur est modifiée par différents facteurs, notamment par le pH, la forme neutre de la molécule étant la plus apte à se dimériser. Dans les solvants organiques, les porphyrines sont, par contre, toujours sous forme monomérique.
Propriétés optiques
La conjugaison du macrocycle des porphyrines laisse prévoir une forte absorption de la lumière dans le domaine proche ultraviolet et visible. Le spectre d’absorption présente de fait une bande autour de 400 nm, dite bande de Soret, qui correspond à la transition de l’état électronique fondamental au second état électronique singulet (S0 → S2). La transition S0 → S1 conduit à une série de bandes dans le rouge (500 à 600 nm) appelées bandes Q. Le spectre correspondant est donné, pour une solution de deutéroporphyrine dans un solvant organique (THF), figure 1.3. Cependant, la forme de ce spectre peut varier de façon très significative lorsque la molécule est en solution aqueuse : à de fortes concentrations, la bande de Soret peut subir une translation de quelques dizaines de nanomètres vers le bleu, accompagnée d’une diminution du coefficient d’extinction (figure 1.4). Ces modifications s’expliquent par la formation de dimères.
Pour de faibles concentrations en porphyrine, (inférieures à 10-7 M), le spectre d’absorption est superposable au spectre d’excitation de fluorescence. Ce spectre est très sensible à l’environnement de la porphyrine. Lorsque la molécule est associée aux protéines plasmatiques, aux lipoprotéines ou incorporée dans des vésicules lipidiques, la bande de Soret est décalée vers le rouge. En effet, plus la molécule se trouve dans un environnement hydrophobe, plus le « red shift » est important. Les bandes Q sont elles aussi modifiées par ces changements d’environnement. Un exemple de spectre d’excitation dans différents environnements est donné chapitre 4 (figure 4.2).
Tout comme le spectre d’absorption, le spectre d’émission de fluorescence est sensible à l’environnement. La bande principale se décale d’une dizaine de nanomètres vers le rouge lorsque la porphyrine est incorporée dans des vésicules lipidiques ou associée à des protéines du sérum ou à des lipoprotéines, comme cela sera montré ultérieurement (figure 4.3).
Enfin, les spectres d’absorption et d’émission sont également modifiés par le pH. Ainsi, à pH acide, une bande d’absorption apparaît autour de 400 nm, ainsi qu’un pic de fluorescence à 592 nm [13]. Les formes mono et dicationiques ont en effet des propriétés optiques sensiblement différentes de la forme neutre. La figure 1.5 montre quelques exemples, à divers pH, de l’émission de la deutéroporphyrine en solution aqueuse. Autour de pH 5, la porphyrine existe essentiellement sous sa forme neutre favorisant l’autoassociation. Les dimères et agrégats n’étant pas fluorescents, l’intensité de l’émission de fluorescence est sensiblement plus faible dans cette zone.
Le spectre d’émission de fluorescence à pH 4,7 présente des bandes caractéristiques à 592, 610, 650 et 670 nm. A pH plus acide, l’intensité de fluorescence à 592 et 650 nm est exaltée. Ces modifications signent la protonation successive des azotes centraux des molécules de porphyrines. Au contraire, à pH plus basique, ce sont les pics à 610 et 670 nm qui prennent de l’importance, correspondant à la prédominance de la forme dianionique monomérique de la molécule.
Enfin, les spectres d’absorption et d’émission sont également modifiés par le pH. Ainsi, à pH acide, une bande d’absorption apparaît autour de 400 nm, ainsi qu’un pic de fluorescence à 592 nm [13]. Les formes mono et dicationiques ont en effet des propriétés optiques sensiblement différentes de la forme neutre. La figure 1.5 montre quelques exemples, à divers pH, de l’émission de la deutéroporphyrine en solution aqueuse. Autour de pH 5, la porphyrine existe essentiellement sous sa forme neutre favorisant l’autoassociation. Les dimères et agrégats n’étant pas fluorescents, l’intensité de l’émission de fluorescence est sensiblement plus faible dans cette zone. Le spectre d’émission de fluorescence à pH 4,7 présente des bandes caractéristiques à 592, 610, 650 et 670 nm. A pH plus acide, l’intensité de fluorescence à 592 et 650 nm est exaltée. Ces modifications signent la protonation successive des azotes centraux des molécules de porphyrines. Au contraire, à pH plus basique, ce sont lespics à 610 et 670 nm qui prennent de l’importance, correspondant à la prédominance de la forme dianionique monomérique de la molécule.
Les bicouches lipidiques et liposomes
Les principales propriétés des lipides et des vésicules lipidiques sont aujourd’hui bien connues et ont été l’objet de nombreuses publications comme [93] et [101] par exemple.
Propriétés physico-chimiques des phospholipides
Les phospholipides dérivent du glycérol par estérification des groupements hydroxyles en position 1 et 2. Ce sont des molécules amphiphiles constituées d’une tête polaire et de chaînes aliphatiques hydrophobes (figure 1.7). Les plus abondants ont des chaînes hydrocarbonées ayant un nombre pair de carbones compris entre 14 et 22. Parmi les phospholipides des membranes plasmiques, les phosphatidylcholines sont prépondérantes.
La phase gel, au-dessous de Tc, correspond à une disposition ordonnée des chaînes en conformation trans. Entre elles, des interactions de van der Waals maintiennent un empilement maximum, limitant les mouvements moléculaires à des vibrations et des mouvements de basses fréquences. Au-dessus de Tc, les lipides sont en phase cristal liquide où les mouvements des chaînes aliphatiques sont beaucoup plus rapides et de plus grande amplitude. De nombreuses conformations gauche apparaissent. Les molécules lipidiques peuvent aussi se déplacer dans la bicouche par des mouvements de rotation autour de leur grand axe et de diffusion latérale. Les coefficients de diffusions expérimentaux sont de l’ordre de 10-12 m 2 /s [102]. La fréquence d’échange est donc de 107 par seconde [93]. Les lipides peuvent également passer d’un feuillet à l’autre de la bicouche. Ce mouvement, appelé flip-flop, nécessite que la tête polaire du lipide passe par la partie hydrophobe de la bicouche. Son temps caractéristique est généralement de plusieurs heures, voire de plusieurs jours. Cette mobilité transversale limitée participe du maintien, dans les membranes biologiques, de l’asymétrie des constituants membranaires. Tous ces mouvements peuvent êtres mis en évidence en utilisant des sondes lipidiques fluorescences comme le 1-oeoyl-2-[12-[(7-nitro-1,2,3- benzoxadiazol-4-yl) amino] dodecanoyl] phosphatidylcholine (NBD-PC) [103].
Les vésicules sont classées en fonction de leur taille et du nombre de compartiments qu’elles comportent. Les vésicules multilamellaires (MLV) présentent plusieurs compartiments aqueux concentriques et peuvent mesurer jusqu’à 1 à 2µm de diamètre. Ces MLV peuvent être obtenues très facilement, par dispersion dans une solution tamponnée. Les grosses vésicules unilamellaires (LUV) sont constituées d’une seule bicouche lipidique et d’un seul compartiment aqueux et mesurent 0,1 à 1µm de diamètre. Les SUV, petites vésicules monolamellaires, que nous avons utilisées pour ce travail, ont un diamètre de 25 à 50 nm. Ces vésicules peuvent être préparées selon différentes méthodes (sonication, évaporation d’une solution éthanolique, évaporation en phase inverse ou, comme ce fut notre cas, par extrusion, voir chapitre « Matériel et méthodes »).
Les transports membranaires
Les vésicules lipidiques sont utilisées comme modèles de la partie lipidique des membranes biologiques pour étudier les propriétés membranaires, les fonctions des protéines membranaires et les interactions molécules-membranes. In vivo, le passage membranaire d’une molécule peut se faire, nous l’avons vu, par diffusion passive, diffusion facilité, transport actif ou endocytose. Dans le cas d’une vésicule qui ne contient ni transporteurs ni pompes à protons, seuls les phénomènes de diffusion peuvent être observés. Trois étapes décrivent le processus (figure 1.9) : l’entrée dans la phase lipidique (a), la traversée de la zone apolaire (b) et enfin la sortie de la membrane (c). Selon la nature de la molécule et les conditions physicochimiques du système, chacune de ces étapes peut devenir limitante. Dans le cas des porphyrines amphiphiles carboxyliques, la traversée du cœur lipidique (b) peut être facilitée, dans un sens donné, par un gradient de pH entre les deux compartiments aqueux délimités par la bicouche. Le franchissement des interfaces pourra être caractérisé par les constantes cinétiques d’entrée (kon) et de sortie (koff).
Les lipoprotéines de basse densité
Structure
Les LDL sont de grosses particules d’environ 2 500 000 Da constituées de phospholipides, de cholestérol (éventuellement sous forme estérifiée), de triglycérides et d’une apoprotéine B100 qui comporte, entre autre, 37 tryptophanes et 151 tyrosines [104]. Un modèle consensus a récemment été proposé [105] : une Propriétés de base des systèmes étudiés monocouche de phospholipides et de cholestérol entoure un noyau de lipide constitué de triglycérides et d’esters de cholestérol. L’apoprotéine, organisée autour de la particule, est constituée par une alternance de feuillets β et d’hélices α (NH3-βα1-α2- β2-α3-COOH). Les feuillets forment des ponts avec des lipides associés de façon irréversible, impliquant probablement du cholestérol en phase smectique∗ . Leshélices α sont associées aux phospholipides extérieurs. Hormis le domaine globulaire N-terminal, homologue à la lipovitelline, la majeure partie de l’apoprotéine se situe dans un environnement hydrophobe. Bien que la forme exacte des LDL soit encore discutée, ils sont le plus souvent considérés comme des particules sphériques de 22 nm de diamètre [105, 106]. Il faut cependant noter que des observations par microscopie cryoélectronique ont suggéré une forme plutôt discoïdale, d’un diamètre d’environ 215 Å et d’une épaisseur de 120 Å, voir même ellipsoïde (250 Å × 210 Å × 175 Å) [107, 108]. Une tentative de représentation a déjà été donnée en figure P.6.
Propriétés spectrales
En les excitant à 280 nm, les LDL fluorescent. Leur spectre d’émission de fluorescence intrinsèque présente une bande autour de 330 nm, due aux résidus tyrosine et tryptophane (figure 1.10). A pH neutre, les maximums d’absorption de ces résidus sont respectivement d’environ 275 nm et 280 nm. Ils sont donc généralement excités simultanément. Cependant, le maximum d’émission des tyrosines se trouvant à 305 nm, leur énergie de fluorescence est transmise aux tryptophanes. C’est donc l’émission de fluorescence des tryptophanes qui est observée. Cette dernière est très sensible à la nature de l’environnement. Dans l’eau, le maximum d’émission est à 350 nm. Dans l’éthanol, la bande se décale vers 340 nm et, dans un solvant totalement apolaire comme le dioxanne, elle est à 325 nm. Ce shift s’accompagne d’une exaltation de fluorescence, le rendement étant de 0,14 dans l’eau, de 0,2 dans l’éthanol et de 0,3 dans le dioxanne. Les tryptophanes des LDL ont un maximum d’émission autour de 330 nm. Ils sont donc dans un environnement hydrophobe. Comme nous venons de le mentionner, l’apoprotéine se trouve en effet directement en contact avec la surface lipidique de ces particules [105]. D’ores et déjà, nous pouvons remarquer que la longueur d’onde d’émission des LDL est proche du maximum d’absorption des porphyrines. Dans des conditions de proximité et d’orientation favorables, un transfert de la fluorescence des LDL vers la porphyrine devrait donc être possible, permettant de mettre en évidence l’interaction du fluorophore avec (ou à proximité de) l’apoprotéine.