Les métabolites spécialisés des plantes
L’ évolution du règne végétal comprend de nombreux exemples démontrant la forte capacité des plantes à s’adapter aux différents milieux présents sur notre Terre. En effet, la sélection naturelle a poussé les végétaux à développer des stratégies de plus en plus complexes afin d’augmenter leur valeur adaptative face à leur environnement. La production de métabolites spécialisés par plusieurs espèces végétales représente sans doute l’une des stratégies adaptatives la plus importante. Ceux-ci permettent aux plantes d’interagir adéquatement avec leur environnement biotique et abiotique. S’étant adapté à pratiquement tous les types d’écosystèmes, le règne végétal présente une immense diversité de métabolites spécialisés. En fonction de leur structure chimique de base, ceuxci sont catégorisés en trois principales classes; les alcaloïdes, les terpénoïdes et les composés phénoliques. Pour la plante elle-même, ces composés peuvent servir à attirer des pollinisateurs ou des disperseurs de graines, à se défendre contre des prédateurs et pathogènes (insectes, herbivores, microbes), à améliorer leur ··Fitness lors de stress abiotique et même à communiquer avec les autres plantes environnantes. Certains métabolites spécialisés peuvent être utilisés comme composés allopathiques contre des compétiteurs.
Aujourd’hui, plus de 50000 métabolites spécialisés des plantes (MSP) ont été identifiés (Wink 2010, Kroymann 2011). En réalité, ce nombre représente qu’une infime proportion de ce qui se trouve dans la nature. Possédant une grande variété d’actions biologiques, ces molécules chimiques végétales sont depuis longtemps exploitées par l ‘homme à titre de médicament, d’aliment, d’arôme, d’insecticide, de pesticide, etc. (Wurtzel and Kutchan 2016). Bien que ces biomolécules représentent un potentiel économique important pour plusieurs secteurs d’activité, la majeure partie des molécules végétales spécialisées étant commercialisées, sont obtenues par extraction à partir de la plante productrice ou par chimie de synthèse.
La principale limitation associée à l’extraction à partir de la plante est le faible niveau de rendement. De plus, en fonction de chaque espèce, les métabolites spécialisés peuvent être produits en différentes quantités selon les différentes parties de la plante (racine, feuille, tige, graine, etc.), selon les différents stades de croissance (germination, plantule, plante mature, etc.) et finalement selon différentes conditions de stress biotique ou abiotique (Delgoda and Murray 2017). En considérant ces nombreuses variables, il peut devenir difficile d’obtenir en quantité suffisante la molécule d’ intérêt. De plus, les étapes de purification subséquentes peuvent parfois être laborieuses et coûteuses.
Finalement, la difficulté ou même l’impossibilité de cultiver à grande échelle certaines espèces de plantes limite encore une fois la gamme de métabolites d’intérêt pouvant être obtenus et donc commercialisés (Liu, Ding et al. 2017). Parallèlement à leur demande croissante, les enjeux reliés aux changements climatiques auront un impact majeur au cours des prochaines années sur la sécurité d’approvisionnement de plusieurs MSP.
D’un autre côté, étant donné la grande complexité chimique de plusieurs de ces MSP, leur synthèse chimique peut s’avérer complexe puisqu’elle nécessite des catalyseurs parfois toxiques, des solvants polluants et des intermédiaires coûteux. Dans tous les cas, bien que le règne végétal représente un vaste répertoire de molécules à haute valeur pharmaceutique ou biotechnologique, seulement un très faible pourcentage de ceux-ci sont commercialement disponible. Face à ces limitations, la production de MSP hétérologue à partir de microorganismes semble représenter une alternative attrayante tant au niveau économique qu’écologique afin d’accroître le potentiel de MSP pouvant être commercialisé. Grâce aux récents progrès qu’a connus la biologie synthétique, la possibilité de reconstruire, étape par étape, une voie de biosynthèse d’un MSP dans un microorganisme hôte est de plus en plus accessible.
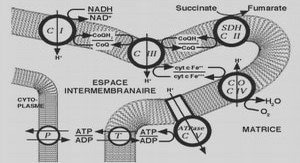
Jusqu’à maintenant, la majeure partie des recherches en biologie synthétique ont été axées sur les organismes modèles bien connus tels que la bactérie Escherichia coli et la levure Saccharomyces cerevisiae (Du, Shao et al. 2011). En effet, à partir de ces deux microorganismes, plusieurs chercheurs ont démontré de bons résultats pour produire quelques composés végétaux hétérologues tels que des terpénoïdes (Ajikumar, Xiao et al. 2010, Ding, Yan et al. 2014) , des alcaloïdes (Hawkins and Smolke 2008, Nakagawa, Minami et al. 2011, Trenchard and Smolke 2015, Diamond and Desgagné-Penix 2016), des polyphénols (Lim, Fowler et al. 2011) et des acides gras végétaux (Zhang, Li et al. 20 Il). Bien que plusieurs ressources génétiques soient disponibles pour la manipulation d’E. coli et de S. cerevisiae, l’utilisation de ces microorganismes pour l’expression de gènes de végétaux présente certains obstacles. Tout d’abord, chez la majorité des eucaryotes, la glycosylation, la phosphorylation ainsi que la formation de ponts disulfure jouent un rôle crucial pour le bon repliement, la stabilité, la solubilité et finalement l’activité biologique des protéines (Walsh and Jefferis 2006). Les différences de certains de ces mécanismes post-traductionnels (PTM) chez E. coli représente l’une des principales limitation pour l’expression de protéines hétérologues actives d’origine végétale (Jenkins 2007). De la même manière, l’absence chez E. coli de l’enzyme cytochrome p450 réductase qui est impliquée dans plusieurs voies de biosynthèse de MSP, peut engendrer un frein pour la reconstruction d’une voie métabolique végétale hétérologue (Du, Shao et al. 2011). Concernant la levure S. cerevisiae, bien que ce modèle eucaryote soit capable de modification post-traductionnel (PTM), la différence de patron de glycosylation avec celui des cellules végétales peut significativement altérer l’activité de l’enzyme d’intérêt in vivo et donc affecter le flux de la voie métabolique insérée (Brooks 2006). De plus, sachant que les différentes étapes enzymatiques d’une voie de MSP surviennent dans différents compartiments cellulaires propres à la cellule végétale, l’absence de plastes chez ces deux organismes modèles peut donc empêcher certaines étapes enzymatiques de se dérouler correctement (Heinig, Gutensohn et al. 2013). Dans le même ordre d’idée, plusieurs MSP sont, lorsqu’ils s’accumulent, toxiques pour la cellule. L’absence de compartiment de stockage telle que la vacuole peut limiter le rendement de production ainsi que la viabilité du système de production hétérologue. Finalement, l’absence de précurseurs ou d’intermédiaire chez l’organisme hôte peut grandement complexifier les étapes de reconstruction d’une voie métabolique hétérologue. Dans cet ordre d’idée, plus on s’adresse aux étapes initiales d’une voie métabolique, plus les chances d’engendrer de la compétition entre différentes voies métaboliques pour un même intermédiaire sont élevées (Xu, Bhan et al. 2013). En considérant ces limitations, il semble évident que l’exploitation d’un organisme modèle végétal phylogénétiquement rapproché des plantes supérieures et pouvant être cultivé à large échelle, permettrait d’accroître la gamme de biomolécules végétales pouvant être produites commercialement. C’est avec cette vision que depuis quelques années, plusieurs chercheurs s’intéressent au potentiel des microalgues comme nouvelle plateforme de production en biologie synthétique (Le6nBafiares, Gonzalez Ballester et al. 2004, Gimpel, Henriquez et al. 2015, Slattery, Diamond et al. 2018).
CHAPITRE 1 INTRODUCTION |