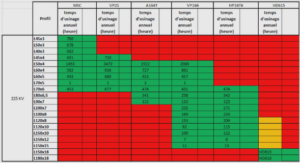
Génération de la diversité
La production d’un nombre très important d’anticorps différents au niveau de leur séquence dans les régions variables est nécessaire pour la reconnaissance de l’immense variété des antigènes du soi et du non-soi (Tonegawa, 1983)(Dudley et al., 2005). Ceci se réalise par des processus de ré-arrangement ou recombinaison de séquences génomiques portant sur les gènes des chaînes lourdes et les gènes des chaînes légères.
Les gènes codant pour les chaînes lourdes sont situés dans le chromosome 14. Ils sont constitués de loci pour les régions V (pour variable) > à 1000, D (pour diversité) > à 15, J (pour jonctionnel)> à 5 et C (pour constante) : μ, δ, γ1, γ2, γ3, γ4, ε, α1, α2) correspondant respectivement aux 9 isotypes de chaînes lourdes. Il existe des exons codant pour chaque domaine constant des chaînes lourdes.
Figure 4. Structure des gènes codant pour les chaînes lourdes
Les recombinaisons sont réalisées grâce aux enzymes Rag (recombinases) Rag1 et Rag2, aussi impliquées dans la génération de la diversité des TCR (Schroeder and Cavacini, 2010). En l’absence de Rag, il existe une déficience en LyT et en LyB par absence de formation des TCR et des BCR. Les étape des processus pour la recombinaison au niveau V-J sont illustrées dans la figure ci-dessous.
Figure 5. Étapes de la recombinaison V-J
Les processus de recombinaison ne sont pas parfaits, sous l’influence de l’enzyme TdT, qui augmente ainsi la diversité. Après les étape de clivage, l’ADN ligase permet la jonction des loci V et J.
Les étape suivantes illustrées dans la figure se succèdent pour aboutir au transcrit primaire qui va être traduit en chaîne lourde.
Figure 6. Étapes de réarrangement des gènes codant pour les chaînes lourdes
Cette organisation génomique permet le mécanisme de commutation de classe, c.a.d. la production, dans un premier temps, des IgM et ensuite, par des mécanismes de recombinaison, des Ig d’autres classes.
Du fait de l’excision de la partie codant la partie constante des IgM, un Ly B produisant par exemple des IgG, ne peut plus synthétiser des IgM (Honjo, 1983).
Concernant les chaînes légères, les processus sont similaires, mais il n’y a pas de régions D et pas de phénomène de commutation de classe.
Les gènes codant les chaînes K se localisent sur le chromosome 2. Deux groupes de gènes codent pour la partie variable de la chaîne κ : Les gènes V qui contiennent 76 gènes et les gènes J contenant 5 gènes. Au cours de la maturation de lymphocytes B, les gènes de la partie variable subissent un réarrangement (sélection et combinaison aléatoire des gènes de la partie variable) avec l’émergence d’un seul gène codant pour la partie variable qui va se combiner au gène codant pour la partie constante (Zachau, 1989).
Les gènes codant pour les chaînes L sont localisés sur le chromosome 22. Les parties variables sont codées par 35 gènes V et 6 gènes J.
Les parties constantes sont codées par un seul gène, il existe un locus les K et plusieurs loci pour les L (Kawasaki et al., 1997).
Figure 7. Structure génomique des gènes des chaînes légères
Une fois les chaînes lourdes et légères synthétisées, leur association va augmenter la diversité du fait du partage du site anticorps. Enfin, lors des réponses immunitaires à un antigène, les LyB vont subir des mutations ponctuelles à l’origine de modification permettant l’augmentation d’affinité pour cet antigène.
La première étape porte sur le ré-arrangement des gènes qui codent pour la chaîne lourde sur un des chromosomes, d’origine maternel ou paternel. Si ce processus échoue, il est réalisé sur l’autre chromosome. La deuxième étape porte ensuite sur le ré-arrangement de la chaîne légère pour aboutir à un LyB mature, portant les Ig de surface. La synthèse des Ig commence à partir du stade pré-B mais elles restent intra-cytoplasmiques. C’est seulement lorsque les Ig sont exportées à la surface du lymphocyte sous forme d’Ig membranaire que le LyB est mature. C’est une synthèse qui se fait indépendamment d’un stimulus antigénique.
Au cours de sa différenciation, le LyB produit d’abord des Ig membranaires à sa surface, puis des Ig sécrétées après maturation en plasmocyte. Cette étape de maturation de passage de la production de formes membranaires d’Ig par les LyB à la synthèse de formes solubles par les plasmocytes différenciés se fait par un mécanisme d’épissage alternatif. Cette étape est liée à une activation (dans les organes lymphoïdes secondaires) du LyB, liée à la rencontre avec l’Ag, et la coopération avec des LyT helpers. Les LyB synthétisent d’abord des IgM et des IgD et produiront ensuite une autre classe d’Ig (IgG, IgE ou IgA) par le mécanisme de commutation de classe ou « switch » d’isotype (Bankovich et al., 2007)(Geisberger et al., 2006).