Les sites d’observation
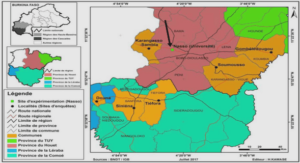
En Guyane française, les sites d’étude sont au nombre de trois (Figure 3) . Il s’agit du Camp des Nouragues, du bassin du Sinnamary ou site de Petit Saut et de la Piste de St. Elie. Les observations qualitatives ont été réalisées indépendamment sur ces trois sites ainsi que sur quelques autres visités ponctuellement. Les mesures quantitatives ont été, en revanche, intégralement réalisées à partir de parcelles établies sur le site de la Piste de St. Elie pour des raisons d’accessibilité, de pérennité et de continuité sur une région déjà bien connue. Ces trois sites ont été utilisés différemment selon les caractéristiques de chacun.
Le site de Petit Saut constitue la référence pour toutes les observations concernant les arbres de la canopée. La mise en eau du barrage de petit Saut, le 6 janvier 1994, a autorisé un accès aisé et progressif aux différentes hauteurs de la forêt mais c’est surtout après le remplissage de la retenue à une cote de 30 m environ, en juillet 1994, que l’observation des grands arbres, depuis un bateau, s’est affranchie des erreurs de parallaxes dues, d’ordinaire, à la vision depuis le sol. Le site des Nouragues est venu en complément des observations de Petit Saut. L’accès à la canopée est permis par quatre passerelles consécutives mesurant au total 105 m. En dehors des arbres traversés par ce « layon » de canopée, la structure verticale sous-jacente a pu être observée à partir de cordes fixées aux passerelles et déroulées jusqu’au sol. Depuis ces axes de circulation, la descente à l’aide du matériel utilisé en spéléologie a permis des observations à tous les niveaux depuis la canopée jusqu’au sol. Les arbres ont été observés, au camp des Nouragues durant deux missions de trois semaines (août 1994 et mars 1995) et à Petit Saut en conjugaison du travail portant sur la reconstruction structurale du massif forestier après la mise en eau effectué dans le cadre d’une convention EDF-ORSTOM (Convention, N° 166, commande GP 7530, avenant n° 1) et ayant donné lieu à trois rapports intermédiaires et un rapport final (Birnbaum, 1996) et la participation à trois journées scientifiques sur l’environnement de Petit Saut (actes des journées de l’environnement, 28-29 mars 1994; 7-8 novembre 1994; 21-22 juin 1995). Les résultats acquis sur Petit Saut ne seront pas repris en détail dans ce travail, les références bibliographiques précitées invitent à s’y référer pour des informations plus précises.
Le site de mesure la Piste de St. Elie
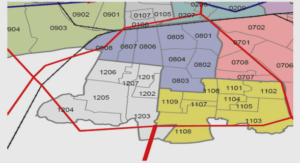
Les parcelles de la Piste de St. Elie, au nombre de trois, se situent au centre du dispositif ECEREX (ECologie, ERosion, EXpérimentation, Figure 4) mis en place en 1977 par le CIRAD (Sarrailh, 1984). Depuis sa création, de nombreux travaux scientifiques y ont eu lieu concernant, entre autres, la pédologie (Boulet et al., 1979), la production de litière (Puig, 1979), la floraison et fructification (Sabatier, 1983), la dissémination (Charles- Dominique et al., 1981), la régénération (de Foresta, 1981 ; de Foresta et al., 1984), la dynamique des chablis (Riera, 1983) , la végétation secondaire (de Foresta et Prévost, 1986), la diversité floristique (Prévost et Sabatier, 1996)… La liste complète des études ayant abouti à une publication est recensée par M-F. Prévost, elle comportait, en 1989, 267 références pour 123 auteurs.
Description de la parcelle 5B-10B de forêt primaire
La topographie de la parcelle est révélatrice de l’hétérogénéité du cadre géographique de l’étude (Figure 7). La carte de la diversité floristique est extraite des données de Prévost et Sabatier obtenues sur la parcelle de 10 ha. Elle est calculée sur des fenêtres mobiles de taille croissante qui balaient la parcelle avec un pas de décalage de 0,01 ha. A chaque pas de balayage, l’indice de Mac Intosh est calculé (Prévost et Sabatier, 1996). La représentation du maillage permet d’analyser visuellement l’évolution de la diversité spécifique des arbres de DBH supérieur à 10 cm le long de la parcelle. La description topographique servira de support à l’énoncé des différents milieux ou paysages forestiers de la forêt primaire. D’est en ouest selon l’axe Y de la parcelle, on distingue 8 séquences (ou topotypes) successives :
De 0 à 70 m (Tp1), la pente moyenne est inférieure à 7 %, constituant la fin d’un méplat.
Le drainage est libre. La structure de cette partie de forêt paraît homogène, les arbres sont grands, le sous-bois est dégagé et l’absence de lianes révèle un état sylvigénétique stable, sans chablis récent (Caballé, comm. pers.). Ponctuellement, les palmiers acaules2 et plus particulièrement Scheelea maripensis, constituent des agrégats. La descente vers le basfond est marquée par une augmentation progressive du nombre des épiphytes et en particulier Philodendron insigne qui forme des manchons continus depuis 8-10 m du sol jusqu’aux premières fourches des arbres et par une diminution de la diversité spécifique des arbres.Entre 70 et 90 m (Tp2), une petite ruisseau (« crique ») traverse un terrain légèrement incurvé qu’il inonde périodiquement et où abondent les Monocotylédones terrestres herbacées (Astrocaryum paramaca, Rapatea paludosa, Heliconia acuminata, H. richardiana, Phenakospermum guyannense, Ischnosiphon obliquus, Thurnia sphaerocephala, Spathiphyllum humboldtii, Spathanthus unilateralis) ou arborescentes (les palmiers Euterpe oleracea, Jessenia bataua) caractéristiques de ce type de « bas-fond ». La structure est caractéristique des bas-fonds, c’est-à-dire marquée par une strate herbacée et une strate arborescente de Monocotylédones et un vide prononcé dans la partie centrale (de Granville, 1978). La diversité spécifique des arbres est parmi la plus basse de toute la parcelle.
Entre 90 et 235 m (Tp3), la pente est importante, de l’ordre de 26 %. Si elle constitue un topotype homogène, cette portion de parcelle est en revanche divisible en deux sous unités caractérisées à la fois par une structure et un état sylvigénétique contrasté et un cortège floristique distinct. La première portion, entre 90 et 150 m c’est-à-dire jusqu’à la fin du 6B, est perturbée par une série de chablis d’âges et de tailles différents. Ces chablis sont orientés dans le sens de la pente et la taille des troncs au sol témoigne d’une forêt anciennement haute. En janvier 1995, un chablis important s’est formé par la chute d’un très grand Dendrobangia boliviana supportant un Coussapoa angustifolia.
La répartition diamétrale selon la topographie de la parcelle 5B-10B
La structure diamétrale du peuplement est généralement utilisée pour mettre en évidence la relation d’équilibre qui lie les jeunes individus aux arbres adultes. Cette relation s’ajuste à une courbe exponentielle décroissante qui indique que le taux de mortalité annuel entre les différentes classes de diamètres varie peu (Rollet, 1969, 1974; Hladik, 1982), les différences dépendant généralement de l’effectif et de la superficie considérés (Riera, 1995). Pour Rollet (1974), l’ajustement des structures diamétrales à une exponentielle est acceptable pour des superficies comprises entre 2 et 16 ha mais plus au-delà de 32 ha. Notre parcelle de 1 ha se situe en deçà de cet intervalle de fiabilité, la droite d’ajustement linéaire est donc volontairement occultée de cette analyse.
Dans la forêt brésilienne proche de Manaus, Guillaumet et Kahn (1982) notent que : « La forêt de plateau, (…), est relativement homogène et marquée par la présence d’un ensemble supérieur d’arbres du présent, bien que peu développé, apparaissant comme un niveau d’émergents. Les pentes dans leur ensemble sont plus hétérogènes. Les bas-fonds se séparent nettement du reste de la toposéquence, mais sont cependant très comparables entre eux. ». Les valeurs d’aire basale sur les hauts de pente et dans les bas-fonds sont plutôt inférieures à celles obtenues sur les plateaux et les pentes. L’aire basale étant corrélée à la densité en arbres de DBH supérieur à 40 cm (Guillaumet et Kahn, Loc. cit.), les hauts de pentes et les bas-fonds seraient caractérisés par une faible densité en arbres de DBH> 40 cm par rapport aux plateaux et pentes (Rollet, 1974 ; Kahn, 1983, 1987; Basnet, 1992; Pelissier, 1995). Torquebiau (1981) montre, que dans la forêt de Los Tuxtlas (Mexique), les plus grands arbres (30-35 m) se concentrent dans les parties topographiques moins « tourmentées » et particulièrement au niveau de la transition entre rupture de pente et plateau, Bonnis (1980) arrive aux mêmes conclusions en forêt de Taï (Côte d’Ivoire). Récemment Pelissier (1995) intègre hauteur et diamètre dans une relation allométrique qu’il compare à la topographie dans la forêt d’Uppangala, en Inde. L’allométrie oppose les plateaux aux fortes pentes, les premiers étant caractérisés par des arbres plus grands.
L’auteur indique que ces différences pourraient s’expliquer à la fois par une composition floristique différente et par une compétition interindividuelle plus intense sur les plateaux qui : « pousserait les individus à s’élever au-dessus de leur concurrents ».
L’hétérogénéité de la parcelle de forêt primaire
La parcelle 5B-10B est composée d’une succession de paysages forestiers très différents.
Les relations entre la topographie, l’état sylvigénétique, la diversité floristique et la structure diamétrale du peuplement montrent à la fois une corrélation et une indépendance des paramètres. Les topotypes de même nature ne sont pas discernables en dehors du basfond caractérisé par des valeurs faibles. Les deux crêtes (Tp4 et Tp8) sont très différentes par leur diversité floristique et leur état sylvigénétique. La toposéquence Tp5 – Tp6 – Tp7 ressort comme une série homogène par les caractéristiques du peuplement alors que Tp5 est, sur le terrain, très différent de Tp7. Le paysage forestier des pentes de même orientation (Tp3, Tp7) est semblable alors que la structure diamétrale ne permet pas de les rapprocher. Enfin, le plateau Tp1 et la crête Tp4 sont quantitativement très semblables bien que leur état sylvigénétique les oppose ; la crête est caractérisée par un chablis, le plateau par sa forêt structurée et sans traces de perturbation…
Il est bien difficile de caractériser une forêt par sa densité ou par le diamètre des arbres. La densité est le principal facteur responsable des différences dans la structure du peuplement alors qu’elle est peu reliée, notamment sur la pente Tp3, à la topographie du site en dehors des bas-fonds. En fonction de la densité, la structure du peuplement évolue, de placette en placette, selon des lois démographiques qu’il est difficile d’associer aux topotypes. La superficie, dénominateur de la densité, détermine la probabilité de rencontrer chaque classe de diamètre mais la signification de cette probabilité évolue, à son tour, avec la superficie.
Une superficie de 1 ha est insuffisante pour caractériser le peuplement des gros arbres sur le site de la Piste de St. Elie (l’arbre le plus gros sur la parcelle de 1 ha, Caryocar glabrum, mesure 84 cm de DBH alors que l’arbre le plus gros sur la parcelle de 10 ha, Pradosia cochlearia, mesure 145 cm de DBH). Au contraire, la parcelle de 1 ha intègre une large hétérogénéité forestière dans la classe des petits diamètres (densité moyenne des arbres de DBH <10 cm calculée sur la parcelle de 1 ha est de 23,1 arbres/ 100 m², pour des valeurs oscillant entre 7 et 45 arbres / 100 m²). La structure du peuplement est, dans ces conditions, très difficilement interprétable puisque la probabilité de rencontrer un arbre de diamètre donné évolue avec la superficie considérée.
Outils
Le télémètre optique
En forêt, les arbres sont mesurés le plus fréquemment par un système optique. La relation géométrique de triangulation entre la position de la mesure, le haut et le bas de l’arbre permet de déduire la hauteur verticale de l’arbre. La mesure angulaire est au coeur de la fiabilité des appareillages, elle permet de mesurer l’angle qui sépare le plan horizontal du sommet et du bas de l’arbre. En forêt tropicale de plaine, si l’on mesure un arbre de 40 m depuis un point situé également à 40 m de son pied sur un sol horizontal, une erreur d’1° de mesure se traduit par une erreur de ± 1,4 m. Cette erreur augmente avec la taille de l’arbre et/ou quand diminue la distance entre l’observateur et la base du tronc. Elle se cumule avec l’erreur induite par les irrégularités du terrain, lorsque la distance au sol est mesurée en déroulant un ruban métrique. Cette dernière atteint 1 m par mètre d’erreur de la mesure au sol, et peut être réduite en utilisant un système optique (dendromètre Blume- Leiss à clisimètre). Ce type d’instrument, qui implique la présence d’une mire sur l’objet mesuré, ne peut pas être utilisé pour mesurer directement la hauteur des d’arbres. En dehors des erreurs liées à la précision de la mesure, ces techniques sont soumises aux difficultés de repérer la cime de l’arbre en forêt. Dès que l’on s’éloigne du tronc, il devient difficile de savoir quelle partie de la couronne est visée et s’il s’agit réellement du point le plus haut de l’arbre. De plus ces techniques sont assez fastidieuses : il faut chercher le meilleur angle de vue, trouver le point le plus éloigné de la couronne offrant la meilleure visibilité, positionner et repérer la mire… Finalement, par défaut technique, la hauteur des arbres n’est pas un paramètre retenu dans les inventaires forestiers.
Méthodes
De la représentation individuelle au profil tridimensionnel
L’objectif de cette analyse est d’acquérir des informations volumétriques sur les arbres qui composent la parcelle. Quel est le volume occupé par la couronne des arbres ? A partir de dessins obtenus sur le terrain et à l’aide de l’outil informatique, une méthode est proposée pour construire une couronne d’arbre en trois dimensions. Chaque arbre est ensuite positionné selon ses coordonnées cartésiennes X et Y et l’ensemble constitue un profil tridimensionnel. La méthode combine l’analyse graphique à celle d’une base de données contenant des informations individuelles (famille, genre, espèce, coordonnées, diamètre…) ou les informations calculées par cette méthode (volume, surface, symétrie…)
Construction tridimensionnelle des arbres
La construction du profil tridimensionnel passe par la reconstitution du volume de la couronne de chacun des arbres. A partir des informations collectées sur le terrain, forcément insuffisantes pour décrire un volume fractal, on ne pourra bien sûr obtenir qu’une approximation de la forme et du volume de la couronne.
L’objectif de la construction tridimensionnelle est de faire coïncider les informations obtenues sur le profil avec celles provenant de la projection de couronne. La description du protocole est volontairement restreinte au principe général de cette construction. Le lecteur désireux de connaître ses fondements géométrique et informatique se reportera à l’annexe
« Protocole de la construction tridimensionnelle des arbres ».
La courbe qui constitue le profil marque la largeur maximale, dans toutes les directions du plan vertical d’observation, de la couronne. Celle qui forme la projection de la couronne au sol marque aussi la largeur maximale de l’ensemble assimilateur, mais dans toutes les directions horizontales et quelle que soit la hauteur.
La projection verticale au sol du profil est un segment de droite dont les extrémités M1 et M2 sont théoriquement sur la projection de la couronne (à noter que le point marquant la position du tronc n’appartient pas nécessairement à ce segment, puisqu’il n’est pas forcément dans le plan de l’observation). M1 et M2, ainsi que les deux points du profil dont ils sont les projections au sol, sont les « points d’ancrage » du profil sur la projection (Figure 13a). Ils servent à ajuster les dimensions du profil (qui ne sont pas mesurées sur le terrain, seules les proportions étant respectées) à celles, mesurées au sol, de la projection. La longueur M1M2 sur le plan de la projection sert de longueur de référence maximale du profil sur la projection.
Une fois ces deux courbes ajustées, la reconstitution du volume de la couronne se fait par étapes :
– Le profil est virtuellement découpé en tranche par des lignes parallèles ou par des lignes qui rejoignent des points équidistantes du sommet de l’arbre (Figure 13b). Le nombre de ligne dépend de la précision recherchée. Les points d’intersection entre ces lignes et le profil donnent deux nouveaux points M1’ et M2′.
– Sur la projection, les lignes sont dans le même plan que le segment M1M2 et le rapport entre leur longueur et celle de ce segment indique l’échelle avec laquelle la courbe de la projection est recalculée. La projection, remise à l’échelle du segment M1’ et M2′ est positionnée par ces points sur le profil.
– Toutes les projections obtenues par approximations successives forment ensemble un volume qui est une représentation de la couronne (Figure 13c).