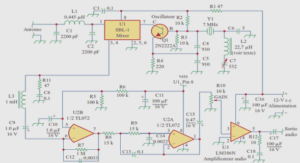
Evaluation de la résistance aux insecticides
Bemisia tabaci populations
A total of 15 field populations of B. tabaci were investigated in this study (Supporting information, Table V-S1 and Figure V-1). They were collected on cultivated plants and weeds all over the island from greenhouses, open fields, field surroundings and non-cultivated areas, from October to December 2017. Field-collected adult whiteflies were transferred to uninfested cotton seedlings (Gossypium hirsutum L. cv. Dora) confined inside insect-proof cages (dimensions: 32.5 × 32.5 × 32.5 cm; mesh size: 650 μm aperture). Each population was established from at least 300 individuals. They were maintained in a climate chamber at 28°C (photoperiod 12:12 h light:dark and relative humidity 60 ± 10%) until they were tested. Susceptible populations of MEAM1 and IO, collected in La Réunion, were used as reference populations when evaluating the insecticide susceptibility of sampled populations. The reference MEAM1 laboratory population was originally sampled in 2001 from cabbage Brassica oleracea L. (Brassicaceae), whereas the IO was sampled on July 2017 from Mexican fireplant Euphorbia heterophylla L. (Euphorbiaceae) and lantana Lantana camara L. (Verbenaceae). Both whitefly species were reared on cotton plants in separate climate chambers, using identical conditions described for the field-collected populations, and were maintained without exposure to insecticides. In addition to living individuals, 30 individuals were sampled at each collection site, preserved in 95% ethanol and held at −20°C for molecular species determination in the laboratory. After DNA extraction, each individual was genotyped using the microsatellite marker Ms145 as a diagnostic locus (Dalmon et al. 2008, Delatte et al. 2011). Allele sizes under 200 base pairs (bp) are specific to the IO species, whereas alleles over 200 bp are only found in the MEAM1 species. The partial 3′ mitochondrial cytochrome oxidase I gene (COI) of 53 individuals, randomly chosen from each of the two species, was sequenced in order to confirm the species identification by pairwise nucleotide comparisons with reference sequences from GenBank. For dose-response leaf-dip bioassays, only unmixed species populations were used. In the case of mixed species populations detected on field-collected individuals, a posteriori molecular species determination was performed on all individuals after each bioassay (IO and MEAM1, i.e. P60 and P64).
Dose-response leaf-dip bioassays
We investigated two of the main classes of insecticides used in La Réunion for the dose– response bioassays: formulated acetamiprid (Supreme® 20 SG, 200 g kg-1 , Certis) and formulated pymetrozine (Plenum 50 WG, 500 g kg-1 , Syngenta Crop Protection), which belong to neonicotinoids and pyridine azomethine derivatives, respectively. Acetamiprid acts as an agonist of nicotinic acetylcholine receptors, inducing hyperactivity and muscle spasms that can lead to death (Jeschke and Nauen 2008). Pymetrozine acts by suppressing stylet penetration so that whiteflies can no longer feed on the plant and die of starvation (Harrewijn and Kayser 1997). Bioassays were performed for both insecticides on each field population (with the exception of populations P35 and P60-IO that were only tested against acetamiprid because of an insufficient number of adults available in the population) using a leaf-dip method adapted from the Insecticide Resistance Action Committee protocol 015 (IRAC; http://www.iraconline.org/methods/). Untreated cotton plants were pot cultivated in a growth chamber (temperature 28°C, photoperiod 12:12 h light:dark and relative humidity 70 ± 10%), away from any pests. About 6 weeks after germination, their leaves were removed and 86 mm diameter discs were cut out from them. From 4.096 and 10 g L −1 stock solutions of acetamiprid and pymetrozine, respectively, two-fold or four-fold serial dilutions of insecticide were made with distilled water. Each cotton leaf disc was dipped for 5 s into one of the dilutions of the insecticide. Natural mortality was assessed with control leaf discs treated with distilled water. Leaves were laid on an agar bed (1%) in Petri dishes (88 mm in diameter), with their adaxial surface downwards. For each field population, adult whiteflies were collected from rearing cages and anaesthetized by cold treatment before being sexed under a Leica MZ6 stereomicroscope (Calvert et al. 2001). Adult female whiteflies were placed on each treated leaf disc. Petri dishes were sealed with a transparent ventilated lid and were randomly distributed into climate chambers at 25°C (photoperiod 12:12 h light:dark). Mortality was recorded after 48 or 96 h of exposure to acetamiprid or pymetrozine, respectively. An insect was considered dead if it showed no sign of movement during 10 s. Each replicate consisted of 15 adult female whiteflies for each different concentration tested. A replicate was discarded if the mortality in the control leaf disc was greater than 15%. Each bioassay was performed with at least five concentrations (plus the control), with three to four replicates each. We did not meet some of these protocol requirements for MEAM1 field-collected population P32 and mixed species populations P60 100 and P64. Although population size was smaller or natural mortalities were higher than expected, we performed analysis of the mortality data.
Dose-response curves analysis
Dose-response curve analysis was carried out in R (R Core Team 2017) using the ‘drc’ package v.3.0.1 (Ritz et al. 2015). The insecticide concentration causing 50% whitefly mortality (lethal concentration, LC50) was estimated using the three-parameter log-normal dose-response model, with unbounded level of natural mortality, similar to the log-logistic model of Finney (Finney 1979). This model calculates 95% confidence limits (CL) of LC50 and the slope of the curve around LC50, and provides the background mortality before increasing insecticide dose has any effect. The effect of population or environment on LC50 was assessed with a deviance analysis (Crawley 2005), comparing the deviance of a model specifying different values for the three parameters (i.e. LC50, slope and lower limit) for each population (or each environment, see Supporting information, Table V-S1) to a model with only one value of LC50 for all populations (or environments). Pairwise comparisons of LC50 were achieved using the ‘drc’ function compParm. Deviance analyses and pairwise comparisons of LC50 for MEAM1 field-collected populations tested with pymetrozine could not be performed due to very high resistance levels among populations, leading to convergence failure. The resistance ratio RR50 was computed by dividing the estimated LC50 value of a given field-collected population for a particular chemical by the corresponding LC50 value for the susceptible laboratory population. As described by Ahmad et al. (2007), resistance was classified as none (RR = 1), very low (RR = 2 – 10), low (RR = 11 – 20), moderate (RR = 21 – 50), high (RR = 51 – 100) and very high (RR > 100). Finally, we tested the relationship between the background mortality and the LC50 to acetamiprid using a linear model and an F-test. The expectation was that resistant populations (with higher LC50) should have lower survival than susceptible populations at sublethal doses if there is a high fitness cost (i.e. causing mortality in the absence of treatment) associated with resistance to acetamiprid.
Life history traits
We further investigated the potential cost of resistance to acetamiprid by recording six fitness-related traits (fecundity, female adult life expectancy, hatching rate, egg-to-adult survival, lifetime reproductive success and female size) for several populations that displayed different levels of resistance. Adult size can be considered as a fitness proxy in females since it is directly correlated with fecundity (Honěk 1993). The cost of pymetrozine resistance was not investigated since it was not possible to determine precisely LC50 values for some MEAM1 populations. Based on the previously described bioassays, we selected seven field populations of the MEAM1 species, in addition to the reference laboratory population. During the following experiments, whitefly individuals were randomly distributed over cotton plants and in climate chambers at 28°C (photoperiod 12:12 h light:dark). Female adult life expectancy and fecundity were measured for ten females per population. To collect newly emerged females, cotton leaves with whitefly pupae were cut and placed in Petri dishes on an agar bed (1%). The next day, newly emerged adults were collected, anaesthetized by cold treatment and sexed. One female and one male (about 1 day old) were placed together in a clip-cage (modified from Millot (2017)) attached on a cotton leaf and cotton plants were then transferred into climate chambers. Three days later, we checked if both male and female were alive and if so the male was removed and the clip-cage (with the female) was moved to a new cotton leaf. The clip-cages were moved every 3 days to another cotton leaf until the death of the female. Eggs laid over each 3-day period were counted. Fecundity was calculated as the sum of eggs laid until death by each female. Female adult life expectancy was the number of days a female lived after emergence, and this observation was made at 3-day time intervals. We monitored egg hatching, egg-to-adult survival and adult emergence on eggs laid on cotton leaves during the first 3 days of this experiment. Leaves were cut 15 days after the start of the experiment and placed on agarose gel in a Petri dish. Every 3 days leaves were observed using a Leica MZ6 stereomicroscope, and all individuals (larvae and adults) were counted and their developmental stages were determined. Emerged adults were repeatedly removed to avoid multiple counting. Therefore, for each female, we measured the number of eggs laid during its first 3 days of life, the number of larvae that hatched and the number of emerged adults. These data were used to estimate hatching rate (number of larvae divided by the number of eggs) and egg-to-adult survival (number of adults divided by the number of eggs). Lifetime reproductive success was estimated by multiplying egg-to-adult survival by the total number of eggs laid in 102 a lifetime (recorded during the duration spent in clip-cages). Finally, we measured the body size, from the tip of the head to the end of the abdomen, of 24-h-old females (20 per population) under a Leica MZ6 stereomicroscope. All data and scripts used to perform the analyses presented in this paper are available in the zenodo repository http://doi.org/10.5281/zenodo.3266422 and at https://dataverse.cirad.fr/.