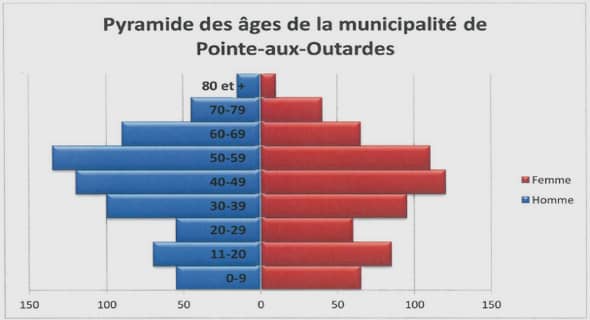
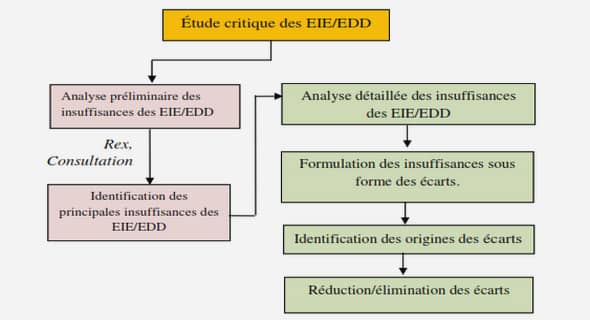
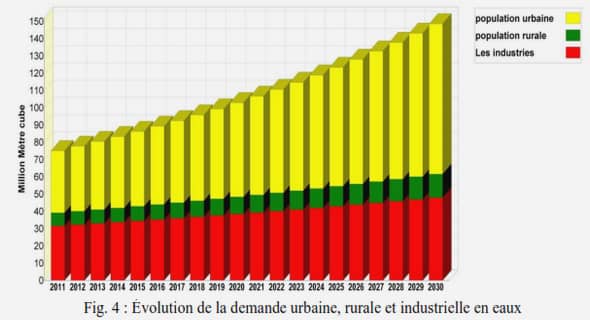
Diversité génétique
Après le nettoyage et l’alignement des séquences, notre jeu de données se trouve constitué à la fin de 38 séquences du gène cytochrome b d’une longueur de 393 nucléotides. De ces 38 séquences, 21 sont de la forme voilière de C. maculatus et 17 de la forme non voilière. Afin de confirmer que les séquences sont bien de la même espèce, elles ont subi un blast (voir annexe 4). Les valeurs de la variabilité sont présentées dans le tableau (1). Pour la forme voilière, 6,87% des sites sont variables parmi lesquels 55,55% d’entre eux sont des sites variables en parcimonie (informatifs). Pour la forme voilière, 10,18% des sites sont variables dont 12,5% d’entre eux sont des sites informatifs. Le taux de substitution synonyme est plus important que le taux de substitution non synonyme pour les deux formes. Le tableau (2) présente la liste des haplotypes et le nombre d’individus qui les composent. On observe que 8 haplotypes sont exclusifs à la forme voilière et les autres à la forme non voilière. Aucun haplotype présentant les deux formes n’est noté. Le tableau (4) présente la moyenne de la composition nucléotidique de la forme voilière et de la forme non voilière de C. maculatus. Ce tableau révèle l’existence d’une différence notable au niveau des moyennes de thymine et cytosine entre la forme voilière et la forme non voilière.
Les changements d’acide aminé sont répertoriés dans le tableau (5) selon les haplotypes. De ce tableau, les mutations observées sont spécifiques à une forme ou une autre. Cependant ces mutations ne sont pas suffisamment réparties dans une forme ou une autre pour en être caractéristique. Le tableau (6) présente les FST obtenus avec en italique les FST intra population et en gras le FST inter population. Avec un FST au sein de chaque population égale à 0,269 et 0,277 pour respectivement la forme non voilière et la forme voilière de C. maculatus, on note une différenciation très importante entre les individus au sein de chacune de ces populations. Le FST entre population montre également une différenciation très importante. Pour les FST intra population de même que pour celui inter population, on note une p-value nulle et donc hautement significative. intra population en italique et la distance inter population en gras. Ces distances sont faibles dans l’ensemble même si l’on observe une distance inter population légèrement plus grande.
Les valeurs des indices d’évolution démographique sont présentées dans le tableau (9). La forme non voilière présente un D de Tajima négatif et un Fs de Fu positif cependant ces indices possèdent une p-value non significative signe d’une population en expansion modérée. La forme voilière, avec un D de Tajima positif et un Fs de Fu négatif possédant des p-values population constante révèlent un tracé différant pour la forme voilière et la forme non voilière de C. maculatus. Ces graphiques (figures 15 et 16) et présentent cependant une distribution multimodale signalant une population stable. déviations) et le Rg (indice d’irrégularité) sont présentés dans le tableau (10). Les p-values du SSD et du Rg sont pour les deux formes non significatives. Les valeurs non significatives de SSD montrent que l’écart observé entre distribution des fréquences observées et attendues n’est pas significatif.
Reconstructions phylogénétiques
Les figures représentent les arbres phylogénétiques obtenus selon respectivement les méthodes de Neighbor-Joining, du maximum de vraisemblance et de l’inférence bayésienne. Ces arbres montrent un regroupement des individus respectant la forme voilière ou non voilière de l’insecte. La forme non voilière étant plus proche de notre outgroup Caryedon serratus. Ainsi, à la suite de ce séquençage, 38 séquences d’une longueur de 393 nucléotides sont obtenues. De ces séquences, 17 appartiennent à la forme non voilière et 21 à la forme voilière. Ces séquences présentent des pourcentages de sites conservés élevé (93,13% et 89,82%), ce qui est également observé dans les travaux de Barry et al. (2017). On note que la forme voilière possède plus de site variable que la forme non voilière. Cependant, la forme non voilière compte plus de site variable informatif que la forme voilière. Cette variabilité plus importante dans la forme voilière que dans la forme non voilière se ressent au niveau des haplotypes où l’on observe plus d’haplotypes avec la forme voilières. La répartition de ces haplotypes (tableau 2) montre qu’aucun d’entre eux n’est identifié à la fois dans les deux formes C. maculatus. Autrement dit, tous les haplotypes présentent uniquement une forme ou une autre de C. maculatus. Ceci peut s’expliquer par la présence d’une transition entre T vers C à la position 354 existant uniquement chez les individus de la forme non voilière néotène. Cette mutation synonyme qui semble sans effet direct sur le gène car l’acide aminé formé n’est pas modifié pourrait faire partie des signaux si elle est couplée à d’autre existant dans d’autre gènes entrainant l’apparition de la forme voilière. Dans ce cas présent, on peut penser que la transition T – C à la position 354 serait importante dans le phénomène de méthylation permettant la régulation de l’expression des gènes. Ainsi en changeant le nucléotide T en C méthylé, on aurait une « inhibition » permettant l’apparition de la forme non voilière ce qui concorde avec l’hypothèse de Utida (1972) et Nahdy et al.,(1999). L’importance du phénomène de méthylation dans ce type de dimorphisme est notée par Brisson (2010) dans ces travaux sur le dimorphisme des ailes chez les pucerons. L’existence du phénomène de méthylation dans le cytochrome b est également démontrée dans les travaux de Iacobazzi, et al., (2013).