Croissance pondérale absolue
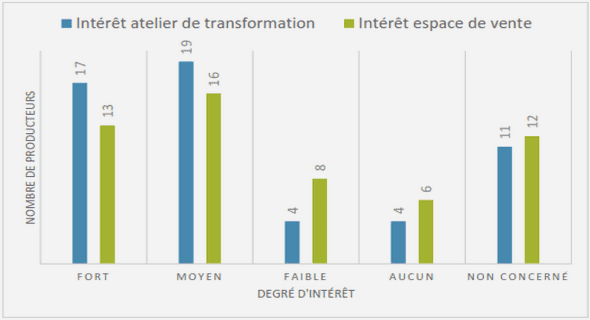
Chez E. encrasicolus et S. pilchardus, l’application de l’équation de croissance pondérale nous a permis de calculer les poids théoriques moyens aux différents âges et de tracer les courbes de croissance pondérale théorique. Chez E. encrasicolus, l’accroissement pondéral est de 5.72 g durant la 1ère année de vie. La croissance en poids est accélérée au cours de la 2ème et la 3ème année avec des gains respectifs de 11 g an-1 et de 8.44 g an-1. A partir de la 4ème année, l’accroissement pondéral des anchois diminue pour atteindre 4.90 g an-1 (Tab. 29, Fig. 39). Chez S. pilchardus, l’accroissement pondéral est important durant sa 1ère année. Elle peut atteindre en moyenne 11,74 g. Dès la 2ème année, la croissance en poids diminue avec un gain pondéral de 8.53 g an-1. Ce taux reste stable au cours de la 3ème année avec 8.12 g an-1, puis il vat diminuer pour atteindre une valeur plus faible de 6.96 g an-1 durant la 4ème année (Tab. 30, Fig. 40). Ceci montre que la croissance pondérale de la sardine est importante durant sa 1ère année de vie. Il est stable au cours de la 2ème et la 3ème année, faible durant la 4ème année.
L’anchois, Engraulis encrasicolus et la sardine, Sardina pilchardus, 2 Téléostéens pélagiques abondants le long du LEA où ils sont capturés toute l’année. Les échantillons que nous avons obtenus mesuraient de 7.8 à 17.6 cm avec une moyenne de 13.79 cm pour l’anchois et de 7.3 à 18.7 cm avec une moyenne de 13.42 cm pour la sardine. Chez les 2 espèces, il n’existe pas de caractère sexuel secondaire apparent qui permet de distinguer les mâles des femelles. Le sexe est déterminé alors après dissection et observation macroscopique des gonades. La proportion des sexes est directement liée à la reproduction, en effet, plus le rapport entre les mâles et les femelles est proche de l’équilibre (1), plus la reproduction comme le montre Legendre (1991) est meilleure. Chez E. encrasicolus, le sex-ratio est globalement constant (SR = 0.98 : 1) et sans différence significative. Les taux de masculinité et de féminité sont presque égaux et proches de 50%. Ce résultat est comparable avec ceux trouvés chez la même espèce aussi bien dans la Méditerranée (Giraldez et Abad, 1995; Sinovčić, 2000; Gaamour et al., 2004; Khemiri et Gaamour, 2009; Morello et Arneri, 2009) que dans l’Atlantique (Millàn, 1999). Cependant, Manzo et al. (2013) montrent que le sex-ratio d’E. encrasicolus des côtes italiennes est en faveur des femelles. Le même résultat a été signalé par Ba (1988) sur l’anchois des côtes mauritaniennes. Des données similaires sont rapportées chez l’anchois du Pérou, Engraulis ringens (Alheit et al., 1984) et celui de la Californie, Engraulis mordax (Klingbeil, 1978; Parrish et al., 1986) où les femelles dominent nettement dans les prises.
La dominance des femelles dans les petites classes de tailles est constatée par l’identification précoce et assez aisée des petits ovaires par rapport aux petits testicules. La supériorité numérique des femelles dans les grandes classes de tailles est signalée par plusieurs auteurs (Giraldez et Abad, 1995; Millàn, 1999; Ouattara et al. 2008). C’est probablement le concours de quelques facteurs dont les plus possibles seraient une plus grande longévité et une croissance plus rapide des femelles, une importante vulnérabilité vis- à-vis des engins de pêche, une mortalité naturelle différente selon le sexe (Garcia et Albaret, 1977; Parrish et al., 1986; Motos et Uriarte, 1991), mais l’hypothèse la plus plausible serait la migration. En effet, le déplacement spatio-temporel des anchois et des sardines affecte différemment toutes les classes de taille (Marchal, 1993). L’analyse des variations mensuelles des 5 stades de maturité sexuelle montre que les gonades d’E. encrasicolus et de S. pilchardus se trouvent à différents stades de maturité au cours de l’année. Pour le même mois, il existe une grande variabilité des fréquences des stades de maturité traduisant le grand décalage aussi bien dans la maturation que lors de l’émission entre les individus de la même population. Chez les 2 espèces, la présence de poissons matures que nous avons observé presque tout le long de l’année avec des pourcentages variables traduit l’étalement de la période de reproduction, ce qui a pour conséquence un fractionnement, comme le montrent Kartas et Quignard (1984), de l’émission des œufs en plusieurs «fournées».
Chez E. encrasicolus, des tailles de 1ère maturité sexuelle similaires à celles trouvées ont été mentionnées en Méditerranée (Fage, 1911; Furnestin, 1945; Arrignon, 1966; Girâldez et Abad, 1995) et dans l’Atlantique Est (Ba 1988; Millàn, 1999; Basilone et al., 2006). En revanche, des tailles supérieures à celles du LEA ont été observées chez la même espèce sur les côtes algéroises (Djabali et al.,1988; Sennai, 2003) et dans le golfe de Gascogne (Duhamel et Masse, 2004). Toutefois des tailles inférieures ont été rencontrées chez l’anchois par Marchal (1991), Leonid et Andrianov (1996), Torstensen et al. (2000), Gaamour et al. (2004), Mehl et al. (2005) et Ouattara et al. (2008).