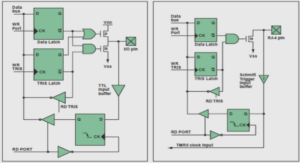
Results and discussion
Wood hygroscopicity
The mean and coefficient of variation of EMC are shown in Table 2.1. The coefficient of variation of EMC values generally increased as relative humidity increased. Sorption at high humidities is mainly affected by capillary forces. Thus, the higher variation in EMC at high humidities could be principally due to variations in the capillary structure among the twenty samples used.
The relationship between water potential and EMC for yellow birch wood is given in Figure 2.1. Previous results obtained for sugar maple wood (Hernández and Bizoň 1994) are also added for comparative purposes. This figure only displays the desorption curve obtained by using either the pressure membrane or the saturated salt solution methods. In all cases, the standard errors of the EMC data do not exceed the symbol size shown. An excellent continuity is apparent between the results obtained by both sorption methods. This confirms the suitability of the pressure membrane method for determining EMC in wood under high relative humidity conditions, which is consistent with numerous earlier reports (Stone and Scallan 1967; Cloutier and Fortin 1991; Hernández and Bizoň 1994).
In this study the desorption was carried out beginning from the fully saturated state, and the curve obtained corresponds to the maximum EMC expected for each humidity condition. The term boundary desorption curve is therefore used to describe this feature. Any desorption curve obtained from a lower initial MC would be located below this boundary desorption curve.
The region between 96% and 100% RH is greatly expanded when using the water potential concept to represent sorption isotherms. This region is very important when studying the wood-water interactions given that it is mainly controlled by the capillary forces and consequently by the micro-structure of wood species. Since wood is a porous material, an important effect that has to be considered in the interpretation of Figure 2.1 is the “ink-bottle effect”. The capillary system of wood consists of cavities interconnected by narrow channels. The variation in dimensions between the different types of cavities connected in series suggests that desorption tends to be governed by a lower water potential, which is determined by the narrower sections of the pores. In contrast, adsorption tends to be governed by a higher water potential, which depends on the larger sections of the pores. Thus, the desorption isotherm will depend on the size of channels connecting the lumina, whereas the adsorption isotherm will depend on the size of these lumina (Fortin 1979).
Figure 2.1 shows that water drainage between full saturation and -100 Jkg−1 ψ was about 35.5% MC. In terms of mass units, this corresponds to a loss of 5.188 g of liquid water. This water would have occupied a volume of about 21.4% within the wood specimen (mean volume of the specimen at 12% EMC was 24.2 cm3). From the discussion above, it appears that this water would have been removed from the larger capillaries, especially the vessel lumina. According to Panshin and de Zeeuw (1980), the proportion of vessel lumina in yellow birch is 21.4% of the total volume. Table 2.1 indicates that at -100 Jkg−1 ψ capillaries with radius larger than 1.44 μm are already empty. Since the radius of vessel elements in birch species is between 30 and 80 μm (Brown et al. 1949), it is apparent that all vessel elements were empty at this stage of desorption.
The boundary desorption curve of yellow birch changes sharply between -100 Jkg−1 and – 700 Jkg−1 ψ, and plateaus between -700 Jkg−1 and -2 000 Jkg−1 ψ (Figure 2.1). Changes in EMC for sugar maple in this region of ψ were more abrupt and the plateau corresponded instead to ψ values between -300 Jkg−1 and -2 000 Jkg−1. This implies that yellow birch presents the least capillaries with a radius larger than 0.48 μm (-300 Jkg−1) compared to sugar maple (Table 2.1). The difference in EMC between the two species at -300 Jkg−1 ψ was about 26%. On the other hand, these species exhibit a similar volume of capillaries with a radius larger than 0.21 μm (-700 Jkg−1 ψ). The results indicate that complete drainage of the fiber cavities would be accomplished at -300 Jkg−1 ψ for sugar maple and at -700 Jkg−1 ψ for yellow birch wood. This drainage would depend on the size of openings between the pit membrane strands connecting fibers. This means that the mean radius of these openings could be larger for sugar maple (0.48 μm) than for yellow birch wood (0.21 μm). Observations undertaken on scanning electronic microscopy (SEM) images (not shown) have corroborated such findings.
The plateau observed between -700 Jkg−1 and -2 000 Jkg−1 ψ for yellow birch indicates that openings controlling the retention and flow of water are scarce within these ψ values. The water remaining in wood would be localized in capillaries having a radius equal to or smaller than about 0.072 μm (Table 2.1). The plateau shown would correspond to the transition between the drainage of the fiber cavities and the drainage of cell walls, and longitudinal and ray parenchyma lumina. As discussed later, an important part of the liquid water remaining below -700 Jkg−1 ψ in yellow birch could be entrapped in the parenchyma cells as noted by Hart (1984).
For softwood species, Fortin (1979) and Tremblay et al. (1996) observed a drainage curve with no intermediate plateau. Aspen wood presented this feature but at higher ψ values (-25 to -100 Jkg-1) than yellow birch and sugar maple (Cloutier and Fortin 1991). Such results confirm that at high humidities, the EMC – ψ relationship is highly dependent on species.
Values of EMC for yellow birch in the range of -2 000 Jkg−1 and -20 750 Jkg−1 ψ values were lower than those reported previously for sugar maple wood (Figure 2.1). This suggests that sugar maple could have more blocked cavities restraining the drainage of the remaining liquid water at these levels of ψ. An analysis of SEM images (not shown) indicated that pit rays in sugar maple were very scarce.
The boundary desorption curves of yellow birch and sugar maple almost join below 76% RH (Figure 2.1). This was expected given that desorption of liquid water at this level of RH is almost achieved. More precisely, Hernández (1983) reported that loss of liquid water for sugar maple will be accomplished at about 63% RH, which corresponds to nearly 14% EMC. It is recognized that bound water desorption is quite similar among different temperate neutral woods.
The FSP value of 34.6% for yellow birch obtained in adsorption over distilled water (Table 2.1) was higher than the expected one estimated by the volumetric shrinkage intersection point method (Figure 2.2). The FSP determined by this method (moisture content at which the extended linear portion of shrinkage-moisture curve intersects the line of zero shrinkage) was about 29.2%. For this estimation, only volumetric shrinkage values obtained between 33% and 76% RH were used. This was done because of the non-linearity of the shrinkage-moisture curve at low moisture contents (Kelsey 1956) and the effect of the hysteresis at saturation on shrinkage at high moisture contents (Hernández and Bizoň 1994). It is clear that the FSP value determined by adsorption over distilled water was overestimated due to a water condensation occurring in the cell cavities and intercellular spaces. Hernández (2006b) has indicated that the adsorption test over distilled water for determining FSP is not applicable for all wood species. Condensation occurred in woods exhibiting a large range of densities and micro-anatomical structures. Consequently, the FSP value of yellow birch estimated by the volumetric shrinkage intersection point method will be used in the discussion that follows.
Wood shrinkage – EMC relationships
The relationships between the EMC and the radial, tangential and volumetric shrinkages of yellow birch are shown in Figure 2.2. This figure indicates that radial, tangential and volumetric shrinkages begin at about 41% EMC, which corresponds to -700 Jkg−1 ψ. In this study, the amount of bound water or actual FSP for yellow birch was estimated to be 29.2%, which indicates that shrinkage started before the FSP was reached. This means that, even at equilibrium, loss of bound water within the cell walls provokes shrinkage of wood before all liquid water has evaporated. For sugar maple wood, Hernández and Bizoň (1994) stated that shrinkage begun at 42.5% EMC even though the FSP of this species is 31%. Stevens (1963) also noted that shrinkage begun well before the FSP for beech wood.
This implies that about 12% MC in liquid form is still retained in the wood when shrinkage of yellow birch starts taking place at -700 Jkg−1 ψ (41.2% – 29.2% EMC). In terms of mass units, this corresponds to 1.7 g of liquid water that would have occupied a volume of about 7% within the wood specimen (mean volume of the specimens at 12% EMC was 24.2 cm3). Given the level of ψ concerned, this water could be entrapped in wood cells interconnected by the smallest capillaries or channels. Hernández and Bizoň (1994) stated that this would correspond to the openings in the membranes of the simple pit pairs located between radial parenchyma cells. The presence of liquid water principally in the rays seems possible, given that these wood elements are considered as the least permeable flow path in hardwoods (Siau 1984; Siau 1995). Such elements should be so impermeable that loss of bound water by diffusion starts taking place in the other more permeable wood tissues. Wheeler (1982) noted that the parenchyma-parenchyma pit membranes are thicker than both the intervessel pit membranes and the fiber-fiber pit membranes, and consequently are less efficient pathways for liquid flow. This entrapped water could in fact principally fill the 10.8% volume of ray tissue reported for yellow birch by Panshin and de Zeeuw (1980). The entrapment of liquid water in ray tissue reported by Hart (1984) for hickory and oak adds further support to this hypothesis. Menon et al. (1987) studied the water location during the drying of Douglas fir and western red cedar using proton magnetic resonance technique. Under unequilibrated conditions, liquid water remained in the ray and tracheid compartments when bound water begins to leave the cell wall at 31% MC. The liquid water was all lost when the MC reached values as low as 9%. According to Skaar (1988), the presence of water-soluble materials in the cavities of parenchyma cells may reduce the humidity at which moisture may condense. Detailed studies of wood structure are needed before the location of entrapped liquid water can be known.