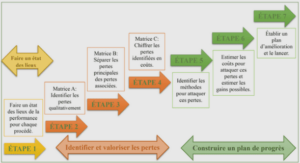
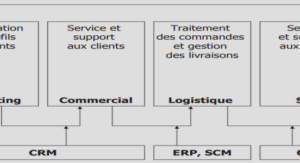
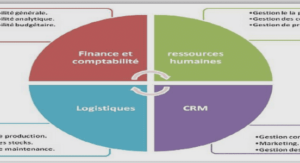
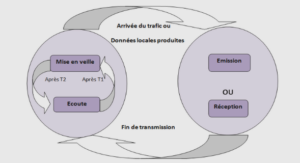
Synthèse bibliographique sur l’acidité des fruits
Fleshy fruit acidity, as measured by titratable acidity and/or pH, is an important component of fruit organoleptic quality (Bugaud et al., 2011; Esti et al., 2002; Harker et al., 2002). Fruit acidity is due to the presence of organic acids, and malic and citric acids are the main acids found in most ripe fruits (Seymour et al., 1993). Understanding the factors that influence the concentration of these acids in fruit cells is thus of primary importance for fruit quality improvement. The predominant organic acid in ripe fruit varies among species. Malic acid is dominant in apple (Yamaki, 1984), loquat (Chen et al., 2009), and pear (Lu et al., 2011), whereas citric acid is dominant in citrus fruits (Yamaki, 1989). In many fruit species, differences in total acidity or in the balance of organic acids among cultivars are also observed, e.g. in loquat (Yang et al., 2011), peach (Etienne et al., 2002), pear (Lu et al., 2011), citrus (Albertini et al., 2006), pineapple (Saradhuldhat and Paull, 2007), apricot (Gurrieri et al., 2001) and banana (Bugaud et al., 2011). The processes involved in the metabolism and accumulation of malic and citric acids in mesocarp cells are under both genetic and environmental control. Transcriptomics (Cercos et al., 2006; Deluc et al., 2007; Etienne et al., 2002), metabolomics (Deluc et al., 2007; Katz et al., 2011), proteomics (Famiani et al., 2005; Katz et al., 2007), and QTLs (Lerceteau- Köhler et al., 2012; Schauer et al., 2006; Xu et al., 2012) studies have helped decipher some of the mechanisms that control acidity, and intervene at cellular level. Many agronomic studies have shown the impacts of cultural practices, including irrigation (Thakur and Singh, 2012; Wu et al., 2002), mineral fertilization (Cummings and Reeves, 1971; Ramesh Kumar and Kumar, 2007; Spironello et al., 2004), thinning (Léchaudel et al., 2005b; Souty et al., 1999; Wu et al., 2002), and environmental factors like temperature (Burdon et al., 2007; Gautier et al., 2005; Wang and Camp, 2000), on fruit acidity, but how they affect malic and citric acid accumulation in the cell is still not clear. In the last few years, process-based simulation models (PBSMs) of fruit have been increasingly used to simulate the metabolic and biophysical aspects of cell behavior (Martre et al., 2011) and appear to be a powerful tool to study genotype x environment interactions (Bertin et al., 2010). Fruit PBSMs of the accumulation of citric acid (Lobit et al., 2003; Wu et al., 2007) and malic acid (Lobit et al., 2006) have been developed to predict citric and malic acid concentrations in the whole fruit during development in peach. The aim of this review is to elucidate the physiological mechanisms that probably control citric and malic acid accumulation during fruit development and their possible regulation by genetic and agro-environmental factors. To this end, the review combines analyses of transcriptomic, metabolomic, and proteomic data related to malic and citric acid metabolism, and also the PBSMs of citric and malic acids. The three first sections describe the cell mechanisms involved in malic and citric acid accumulation and their regulation. The last section deals with the effects of agro-environmental factors (source: sink ratio, mineral fertilization, water supply, and temperature) on citric and malic acid accumulation and the related cell mechanisms they may affect. In this review, the terms “malate” and “citrate”, which usually describe the conjugate base of malic and citric acids, refer to all physiological forms of each compound.
First step in organic acids synthesis:
PEP carboxylation in the cytosol Formation of acidity involves the synthesis of organic acids, mostly malate and citrate, which can be stored in the vacuole in large amounts. As citrate is produced from dicarboxylates (mostly malate) (see following section), the first step in the development of acidity is the synthesis of dicarboxylates, namely malate and OAA. These require fixation of CO2 on a carbon skeleton derived from hexose catabolism (Hardy, 1968; Young and Biale, 1968), which is achieved by the carboxylation of PEP, catalyzed by the phosphoenolpyruvate carboxylase (PEPC). This reaction takes place in the cytosol, since PEP is an intermediate of the glycolysis pathway, and produces OAA, which can then be reduced into malate by the cytosolic NAD-dependant malate dehydrogenase (NAD-cytMDH) (Givan, 1999) or supplied the TCA cycle if replenishment is necessary (Leegood and Walker, 2003) (Fig. I.1). Multiple PEPC isoforms have been detected in fruits and are possibly the result of transcriptomic (Sweetman et al., 2009; Yao et al., 2009) and/or post-translational regulations (Sweetman et al., 2009). PEPC is controlled by both cytosolic pH and malate concentration (Davies, 1986; Lakso and Kliewer, 1975a; Possner et al., 1981) in a way that stabilize the cytosolic pH (Smith and Raven, 1979). In grape berries, transcriptomic analysis (Or et al., 2000; Terrier et al., 2005) and measurement of enzymatic activity (Diakou et al., 2000; Hawker, 1969; Ruffner et al., 1976) pointed to the role of PEPC in malate accumulation throughout fruit development.
Several studies based on analyses of transcriptomic and enzymatic activity suggest that PEPC is not responsible for the difference in malate content between low and high acid peach cultivars (Moing et al., 2000), apple (Yao et al., 2009), and loquat (Chen et al., 2009). NAD-cytMDH catalyzes the reversible conversion of malate into OAA, the most likely direction being the synthesis of malate (Sweetman et al., 2009; Yao et al., 2011). Even if a mitochondrial form is also present in fruit cells (see following section), it has been shown in several fruits that NAD-cytMDH represents 70% to 80% of total NAD-dependant MDH (Abou-Zamzama and Wallace, 1970; Taureilles-Saurel et al., 1995), explaining why total NAD-dependant MDH activity is generally related to malate synthesis in fruits (Chen et al., 2009; Martınez-Esteso et al., 2011; Zhao et al., 2007). Yao et al. (2011) showed that over expression of the apple MdcyMDH gene encoding NAD-cytMDH resulted in an increase in malate, fructose and sucrose content, suggesting its direct involvement in malate synthesis. Over expression of MdcyMDH also resulted in the up regulation of several malate-related genes/enzymes, suggesting an indirect role in malate accumulation.
Organic acids degradation: malate and OAA decarboxylation in the cytosol Loss of acidity implies decarboxylation of carboxylates, which can occur through the conversion of tricarboxylates into dicarboxylates (described later in the review), but also through decarboxylation of the dicarboxylates malate and OAA leading to the degradation of organic acids (Fig. I.1). Decarboxylation of OAA and malate allows the production of PEP and is linked to the activation of gluconeogenesis (Sweetman et al., 2009). Gluconeogenesis is a metabolic pathway that results in the generation of glucose from PEP. It occurs mostly during fruit ripening when sugars accumulate rapidly (Sweetman et al., 2009). In the past few years, proteomics (Katz et al., 2011), transcriptomics and metabolite (Carrari et al., 2006; Deluc et al., 2007; Fait et al., 2008) analyses have provided evidence for a shift from the accumulation of organic acids to sugar synthesis during the final stage of development in several fruit species. PEP can originate from OAA through the activity of phosphoenol carboxykinase (PEPCK) which catalyzes the reversible reaction, the most likely direction being the synthesis of PEP (Leegood and Walker, 2003). This reaction requires a source of OAA that could be supplied by the oxidation of malate by NAD-cytMDH.
This hypothesis is supported by the fact that PEPCK is involved in the dissimilation of malate in the flesh of several fruits (Famiani et al., 2005) and possibly in the lack of malate in low acid apple cultivars (Berüter, 2004). PEP can also originate from the conversion of pyruvate through pyruvate orthophosphate dikinase (PPDK) activity (Sweetman et al., 2009). The pyruvate required for PPDK may be supplied through the carboxylation of malate by cytosolic NADP-dependant malic enzyme (NADP-cytME), which catalyzes a reversible conversion, the most likely direction being the decarboxylation of malate (Sweetman et al., 2009). NADP-cytME appears to be involved in the decrease in malate content during the ripening of several fruit species (Chen et al., 2009; Dilley, 1962; Goodenough et al., 1985; Sweetman et al., 2009). Involvement of NADP-cytME during the early stage of fruit growth differs between species. Thus, in young tomato and apple fruits, NADP-cytME does not appear to play an important role in malate accumulation (Dilley, 1962; Goodenough et al., 1985) whereas in young grape berries the use of a proteomics approach suggested the opposite (Martınez-Esteso et al., 2011). The contribution of NADP-cytME to the lack of malate in ripe pulp of low acid cultivars has been demonstrated in apple (Yao et al., 2009), and loquat (Chen et al., 2009). Studies of different fruit species suggest that NADP-cytME is regulated at the post translational level (Bahrami et al., 2001; Famiani et al., 2000; Yang et al., 2011; Yao et al., 2009) by cytosolic pH and malate concentration, among others (Davies, 1986; Lakso and Kliewer, 1975a; Possner et al., 1981). Decarboxylation of malate and OAA may also be linked to fermentative metabolism as it can occur in ripening fruit if the cytosol becomes too acidic (for review see (Sweetman et al., 2009)).
Introduction Générale |