Régimes d’appariement social et génétique
Les avancées récentes des techniques moléculaires ont complètement révolutionné l’étude des comportements d’appariement animaux en révélant que le régime d’appariement génétique des individus d’une espèce ne pouvait être inféré par de simples observations comportementales des liens sociaux entre les partenaires (Hughes, 1998). Ceci a remis en cause bon nombre d’hypothèses liées à l’adoption de stratégies de reproduction chez les vertébrés. Ainsi, des individus socialement monogames peuvent s’investir dans des événements de reproduction extraconjugaux, ce qui engendre un régime d’appariement génétique de polygamie (Sillero-Zubiri et al., 1996). Au contraire, comme c’est le cas chez beaucoup de canidés de petite taille, le groupe de reproduction peut être caractérisé par la présence d’un mâle pour plusieurs femelles (polygynie sociale), mais d’un seul couple reproducteur (monogamie génétique) (Kleiman, 1977). En fait, tous les régimes d’appariement sociaux peuvent être reliés à n’importe lequel des régimes d’appariement génétique. Les individus font donc face à un double choix dans l’adoption des comportements d’appariement, soit l’obtention d’un ou plusieurs partenaires sociaux mais aussi l’obtention d’un ou plusieurs partenaires génétiques. De plus les partenaires sociaux et génétiques peuvent être identiques ou différents. Il est très important de tenir compte de la distinction entre partenaires sociaux et partenaires génétiques dans l’étude des comportements de reproduction, puisque les paramètres évolutifs façonnant les régimes d’appariement génétiques et les régimes d’appariement sociaux d’une même espèce peuvent être totalement différents. Par exemple, chez le loup d’Abyssinie (Canis simensis), les groupes sociaux sont formés de plusieurs individus proches parents en raison de fortes contraintes de dispersion, mais les femelles s’engagent dans des événement de copulation à l’extérieur du groupe afin de limiter la dépression consanguine (Sillero-Zubiri et al. , 1996).
Paternités extraconjugales: monogamie sociale et polyandrie génétique Les cas de paternités extraconjugales, où des femelles socialement monogames sont fécondées par des mâles hors du couple, sont des exemples fréquents de la disparité entre partenaires sociaux et partenaires génétiques. La liste des espèces génétiquement monogames se révèle de plus en plus courte, et il semble que la monogamie stricte (sociale et génétique) représente un cas rare où un ensemble de contraintes force les individus de chacun des sexes à n’avoir qu’un seul partenaire. Les régimes d’appariement d’un grand nombre d’espèces d’oiseaux, canidés, rongeurs et primates classifiées comme monogames ont ainsi dû être reconsidérés à la lumière des nouvelles connaissances sur les comportements menant à la polyandrie. Ceci a fait naître un vif débat sur la valeur évolutive de tels comportements et sur les avantages que les femelles pourraient retirer de la polyandrie. Les principales hypothèses proposées concernent les bénéfices directs pour la femelle et la qualité, diversité et compatibilité des gènes des jeunes produits (Reynolds, 1996). La polyandrie peut d’abord être avantageuse pour les femelles, par les gains directs qu’elles en retirent de la part des mâles. Ces bénéfices peuvent être sous forme de soins paternels additionnels (Davies et Hatchwell, 1992), d’un approvisionnement en sperme suffisant (Ridley, 1988) ou de dons nutritifs de spermatophores ou de liquide séminal (Zeh et Smith, 1985).
Par contre, chez bien des espèces, ces bénéfices ne peuvent s’appliquer et la présence de bénéfices indirects pour les femelles est requise pour expliquer les comportements polyandres. L’hypothèse des « bons gènes» (Orians, 1969) est prédominante dans l’étude du choix du ou des partenaires chez les femelles, et donc de la polyandrie et des comportements extraconjugaux. Selon cette hypothèse, les femelles s’accouplant avec plus d’un mâle pourraient être en mesure de sélectionner le sperme du mâle de meilleure qualité génétique, c’est-à-dire celui qui transporte les allèles procurant la valeur adaptive la plus élevée. L’effet des bons allèles est additif, c’est-à-dire que les femelles devraient toutes préférer les mêmes mâles, indépendamment de leur propre constitution génétique (Neff et Pitcher, 2005). Ces bons allèles pourraient améliorer, chez les jeunes produits, la survie (e.g. résistance aux maladies), la fertilité (qualité du sperme), la compétitivité (condition, vigueur, etc.) ou l’attractivité pour le sexe opposé. Un allèle pourrait également avoir un effet bénéfique chez l’un des sexes et être neutre ou même négatif chez l’autre (Chippindale et al. , 2001).
Il est également possible que le choix des femelles se fasse sur la base de leur compatibilité génétique avec leur partenaire (Zeh et Zeh, 1996, Tregenza et Wedell, 2000). Une femelle baserait alors son choix de partenaire de reproduction génétique par rapport à sa propre composition génétique, et ainsi, le meilleur mâle pour une femelle ne serait pas nécessairement le meilleur pour une autre. Contrairement à l’hypothèse des « bons gènes », cette hypothèse ne mène donc pas à une sélection de certaines caractéristiques reflétant la qualité génétique du mâle, mais à une sélection des comportements favorisant le choix de partenaires compatibles (Kempenaers, 2007). Un exemple largement discuté de ce type de comportements est l’ évitement de la consanguinité (Tregenza et Wedell, 2002), qui est caractérisé par l’évitement de l’accouplement avec des individus fortement apparentés (Blouin et Blouin, 1988). Ce faisant, les femelles augmenteraient l’ hétérozygotie de leurs jeunes et potentiellement leur valeur adaptative (Foerster et al., 2003), si on prend pour acquis que les deux traits sont liés (Allendorf et Leary, 1986). Il a été démontré que l’hétérozygotie au complexe majeur d’histocompatibilité (CMH), un ensemble de gènes impliqués dans la régulation des réponses immunitaires, pouvait être corrélée positivement à des composantes liées à la valeur adaptative telles que la résistance aux pathogènes et la fertilité (Apanius et al., 1997) et faisait l’objet d’une sélection sexuelle chez plusieurs espèces (Tregenza et Wedell, 2000).
La diversité génétique au sein d’une portée, plutôt que l’hétérozygotie des jeunes, est un bénéfice potentiel pour les femelles ayant été fréquemment proposé pour expliquer la polyandrie. Il a en effet été suggéré que par la polyandrie, les femelles pourraient augmenter la diversité génétique à l’intérieur de leur portée, et ainsi réduire les risques d’échecs reproductifs, et à plus juste titre pour les femelles qui font face à des environnements variables et inconnus (Watson, 1991). Cette hypothèse pourrait s’appliquer particulièrement chez les espèces produisant un grand nombre de jeunes par portée et pour lesquelles le nombre d’événements de reproduction au cours de la vie est restreint. Afin d’évaluer les facteurs ayant mené à l’évolution de la polyandrie génétique, il importe d’utiliser des modèles d’études chez lesquels on retrouve non seulement une grande variabilité dans les régimes d’appariement, mais également dans les paramètres physiologiques et environnementaux pouvant influencer ceux-ci. Plusieurs espèces de canidés regroupent ces caractéristiques et en font donc de bons modèles dans l’étude de l’évolution des paternités extraconjugales.
Discussion
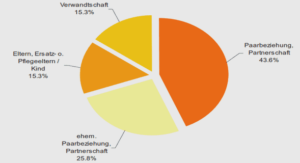
Results from this study suggest that, in the population of arctic foxes from Bylot Island, the dominant social mating system is monogamy with biparental care. This is similar to findings from other arctic fox populations (Macpherson, 1969; Strand et al., 2000). Of aIl dens recorded, only two had an extra adult, in each case a non reproductive female. However, despite their predominantly socially monogamous behaviour, arctic foxes from Bylot Island had high levels of extra-pair paternities; almost the third (31 %) of cubs were born from a male from outside of the social unit, and 31 % of litters with known social father presented EPPs. This is coherent with data from other canid species, in which evidences of extra-pair or extra-group copulations/paternities have been found each time it was investigated (Ethiopian wolf, Canis simensis, Sillero-Zubiri et al. 1996; African wild dog, Lycaon pictus, Girman et al. 1997; swift fox, Vulpes velox, Kitchen et al., 2006; red fox, Vulpes vulpes, Baker et al. 2004; island fox, Urocyon littoralis, Roemer et al. 2001). But the level ofEPP observed in arctic fox es from Bylot Island, ev en if considerably high for a socially monogamous mammal, is stilliower than that of sorne c10sely related canids for which it has been estimated (swift fox: 52%, Kitchen et al., 2006; red fox: 80%, Baker et al., 2004).
It has often been suggested that female mammals may perform extra-pair copulations in order to avoid inbreeding, and to increase their offspring heterozygosity (Tregenza and Wedell, 2002; Foerster et al., 2003). However, in a meta-analysis of 55 bird species, Ackay et al. (2007) found no correlation between the genetic similarity and extra-pair patemity. In the population of arctic foxes ofBylot Island, the fact that at least one case of extra-pair patemity involved incest suggests that inbreeding avoidance cannot explain aU cases ofEPPs. Incest is thought to occur when individuals have incomplete knowledge of relatedness and the costs of avoiding inbreeding are high (Wheelwright et al., 2006). This was most probably not the case in this study, since the male was raised by the female, they were both otherwise socially paired with an unrelated partner, and they were holding a territory located in an area of high resource abundance. Enhanced within-brood genetic diversity rather than offspring heterozygosity seems more likely to have been influential in the evolution ofpolyandrous behaviours in arctic fox es. As proposed by Yasui (1998), if the environmental conditions in which the offspring will develop are predictable, the fitter genes for the next generation are predictable. In those cases, female’s mate choice is expected to go toward males ofhigher quality (genetic quality or heterozygosity, Orians, 1969; Kempenaers, 2007). On the other hand, in a fluctuating environment, females may not be able to predict the fitter genes for the next generation as she may have little cues about future environmental conditions. The increased within-brood genetic diversity ofpolyandrous females may therefore enhance the probability that at least one young survives and reproduces successfully in an unpredictable environment (Jennions and Petrie, 2000). This may be particularly adaptive in environments such as high arctic ecosystems, where harsh and changing conditions prevail.
AVANT-PROPOS |