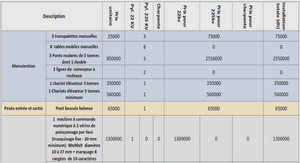
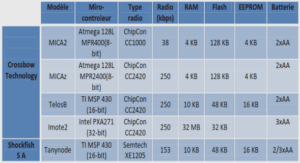
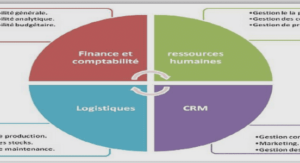
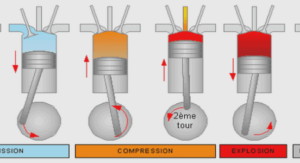
Data Collection
During the summer of 2003, 18 black spruce dominated sites ranging from 50 to 350 years since fire (see Appendix A for a full description) were sampled. A stand initiation map (Bergeron et al., 2004) and an ecoforestry map (Harper et al., 2002) were used to choose sites on a slight incline with fine-textured deposits. Slope and soil textured were both verified at the site, and time since fire was established by either dating of dominant trees, or verification of fire map dates by dating of a few dominant trees. The severity of the last fire on the forest floor was established by determining the position of the uppermost charcoal layer within numerous forest floor profiles. While strictly speaking more fuel may have been consumed in the low severity fires, severity of fire on the forest floor may be most accurately measured by the amount of forest floor not-consumed by fire (Alexander, 1982; Nguyen-Xuan, 2000). If the charcoal was situated at the interface between the forest floor and the mineral soil, the last fire was designated as high severity. If however, the charcoal layer was within the forest floor layer, the last fire was designated as low severity. For further details on the methods used, see Lecomte et al. (2006).
Among the stands sampled, 13 sites were established after high severity fire and five sites after low severity fire. Within each site, five quadrats of 100 m2 were installed, with four nested quadrats of 25 m2 . Within each 25 m2 quadrat, forest floor thickness was determined by measuring the depth to mineral soil in a randomly chosen location, including therefore the entire organic layer. The clay A horizon provided a clear end point to forest floor depth. A densiometer reading was taken (a concave mirror scored with a grid to allow estimation of canopy cover), and the diameter at breast height ( dbh) of all trees greater than 8 em was measured. Basal area per hectare was calculated for all species together, and for trembling aspen alone. Both stand density and canopy openness were included as variables as stand density also includes the effect of the trees on the soil. Covers(%) of Sphagnum spp. and ericaceous spp. (primarily Ledum groenlandicum, and Kalmia angustifolia) were estimated. The 25 m2 quadrats were assumed to be independent, as soil processes tend to vary on the scale of a few meters, therefore analyses were performed on the values for the 25 m2 quadrats.
RESULTS
Forest floor thickness was negatively correlated with fire severity (i.e. thinner forest floors after high severity fires), total basal area, and trembling aspen (PET) basal area, and positively correlated with cover of ericaceous spp. and Sphagnum spp., canopy openness, and time since fire (TSF; Table 1.1 ). The importance of fire severity is visible in the different curves for sites after high and low severity fires (Figure 1.1). After a high severity fire, a 50 year old site had only approximately 17 em of forest floor on average, while in a neighbouring site, 50 years after a fire of low severity there was approximately 40 em of forest floor. As most of the potential factors were auto-correlated, partial correlations were calculated to determine which would be retained for path analysis. TSF, fire severity, cover of Sphagnum spp. and ericaceous spp. were retained, as they had a significant partial correlation, even when the other factors were taken into consideration (Table 1.2). The structure determined by path analysis (Figure 1.2), which was not rejected by d-sep analysis (p= 0.2989; Shipley, 2000), indicated that TSF and fire severity were the two dominant factors on forest floor thickness, with the largest direct effects, 0.606 and -0.295 respectively.
The cover of Sphagnum spp. and ericaceous spp. also had statistically significant direct effects, although with less influence than TSF and fire severity. In fact the majority of the correlation between forest floor thickness and Sphagnum spp. and ericaceous spp. cover is due to shared correlation with TSF and fire severity. In tum, Sphagnum spp. cover was positively influenced by cover of ericaceous spp., and TSF. Cover of ericaceous species was influenced negatively by fire severity and positively by TSF. As fire severity was a dominant factor, the high severity sites were analysed separately. After severe fires, forest floor thickness was highly correlated with the amount of open canopy, cover of Sphagnum spp., cover of ericaceous spp., amount of trembling aspen and TSF, which were all also auto-correlated (Table 1.1). TSF, amount of trembling aspen, cover of Sphagnum spp. and ericaceous spp., and canopy openness all had significant partial correlations when the other factors were accounted for. The structure suggested by path analysis, indicated that only TSF, Sphagnum spp. cover and amount of trembling aspen had direct effects on forest floor thickness (Table 1.2; Figure 1.3). The cover of ericaceous spp. and the amount of open canopy had an indirect effect via Sphagnum spp. cover. The presented model explains 65% of the variability (p<0.0001), as indicated by multiple regression, and the structure was not rejected by a d-sep test (Shipley, 2000) with a p of0.725.
Time since ftre and ftre severity
Time since ftre (TSF) was the most important factor influencing forest floor thickness within both the entire data set and the analyses restricted to the high severity ftre sites. This is only logical, as the matter within the forest floor takes time to accumulate. However the strong relationship with TSF also indicates that on fine-textured soils on the Clay Belt, no additional initiative process is required. This is in contrast to other geographical areas where a non-linear relationship exists, and thick forest floors are developed after the creation of an impermeable layer by an external factor such as the development of permafrost, a chemical change in the soil, or the expansion of neighbouring peatlands (Bonan and Shugart, 1989; Glebov and Korzukhin, 1992; Viereck et al., 1993). From TSF it is possible to calculate the rate of accumulation of forest floor matter. However, the true rate of accumulation in the oldest stands cannot yet be accurately determined, as these sites may be older than indicated by dendrochronological age alone. Carbon 14 tests on similar stands in the same region have indicated that the oldest trees currently found were actually established a long time after the last ftre (Cyr et al., 2005), resulting in an underestimation of stand age.
While a change in stand age would affect the interpretation of the rate of forest floor accumulation and potentially the relative importance of each mechanism, it would not affect the mechanisms investigated here. After TSF, ftre severity was the most important factor in explaining variation in forest floor thickness in the complete data set. This is reflected in the very different forest floor thicknesses in young stands after high and low severity ftres, and in the relationship between forest floor thickness and time (Figure 1.1 ). This may be due to two causes. The intuitive cause is that residual matter left by a low severity ftre gives the forest floor a head start. However, the presence of residual matter may also accelerate the rate of accumulation by affecting the function of factors suggested here, primarily Sphagnum spp. and ericaceous spp. For example, the residual layers of the forest floor left after non-severe ftres may facilitate Sphagnum spp. establishment, and growth (Dymess and Norum, 1983; Purdon et al., 2004). For this reason, a separate analysis was completed for the high severity ftre sites. A similar detailed analysis of the low severity sites was not possible with the data from this study, as too few low severity fires were sampled. Therefore, future studies should focus on understanding the influence of low fire severity and residual matter on the factors influencing the development of forest floors proposed here. This may be of particular importance, as the long fire cycles found in the eastern boreal forests permit the accumulation of a significant forest floor. This may cause a self-perpetuating cycle as forests with a thicker forest floor are moister and lose less organic matter during subsequent fires (Foster, 1985; Kasischke et al., 2000).
Forest floor thickness after high severity fires While TSF was overwhelmingly the most important factor influencing forest floor thickness, there was variation in the thickness of the forest floor within and among sites, particularly at later stages in the chronosequence (Figure 1.1 ). Within this analysis, the factors explaining this variation were the cover of Sphagnum spp., the amount of trembling aspen, the openness of the canopy and the cover of ericaceous spp. The presence of Sphagnum spp. in the understory has frequently been implicated as a key factor in the development of thick forest floors. They are believed to have a negative effect on decomposition through the development of a cold, wet and acidic environment for decomposers (Turetsky, 2003). In addition they also affect decomposition rates through their rapid production of biomass, which has a high C:N ration, and is resistant to decomposition (Hobbie, 1996; Turetsky, 2003).
However, while our results indicate that they are a direct factor in determining forest floor thickness, the magnitude of effect is comparatively weak. The direct negative effect of the presence of trembling aspen on forest floor thickness is inferred to be due to the presence of a high quality litter on the forest floor, which results in higher decomposition rates overall. Legare et al. (2005) found increasing decomposition rates of popsickle sticks, in the forest floor with increasing presence of trembling aspen in the forest composition. In addition to this direct effect, the presence of trembling aspen diminished the cover of Sphagnum spp. This effect is particularly interesting when canopy openness is accounted for and implies that there is a relationship that is not directly related to shade. It has been suggested that deciduous leaves have a negative effect on the growth of forest floor mosses, either through a chemical interaction or through shading (Frego and Carleton, 1995; Saetre et al., 1997). Canopy openness had only an indirect effect on forest floor depth, via Sphagnum spp. cover. The negative interaction between these two factors is surprising, as Sphagnum spp. is generally considered to be shade intolerant, and dependent on full sunlight for maximum growth (Bisbee et al., 2001; Ohlson et al., 2001). However, Sphagnum capillifolium, a highly shade tolerant species (Hayward and Clymo, 1983), dominates in young stands (Fenton and Bergeron, 2006). As a result, total Sphagnum cover does not display the expected relationship with canopy openness. The negative relationship may be due to improved growth of Sphagnum spp. in the less illuminated sites in the older sites, which allows them to avoid desiccation and photoinhibition.
The cover of ericaceous species had only an indirect effect on forest floor depth, via Sphagnum spp. However, in Europe Empetrum nigrum, has been shown to have a strong direct impact on ecosystemic processes (Wardle et al., 1997, 2003) through the production of phenols that retard decomposition (DeLuca et al., 2002). In this study, while Kalmia angustifolia and Ledum groenlandicum litter have been shown to produce phenols (Inderjit and Malik 1997, 2002), their effect on forest floor thickness after high severity fires was solely through their effect on the abundance of Sphagnum spp., and probably not a direct effect on decomposition rate. Ericaceous species, and vascular plants in general, may stimulate Sphagnum spp. growth through a « scaffolding » effect, where the Sphagnum spp. use vascular plants to physically support fast vertical growth (Maimer et al., 2003). The high level of variability in forest floor depth within sites, particularly in the older sites (Figure 1.1A), indicates the importance of very local factors, such as small canopy openings, and Sphagnum spp. and ericaceous spp. cover.
REMERCIEMENTS |