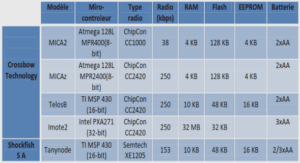
Propriétés chimiques
Le potentiel d’hydrogène (pH) du sol est reconnu pour réguler la biodisponibilité des nutriments pour les plantes (Truog 1948). Lorsque l’on observe des pH faibles, la fertilité du sol sera réduite entrainant une diminution de la productivité primaire de l’écosystème (Li Y. et al. 2017). Pour plusieurs espèces, l’augmentation du pH, jusqu’à une valeur limite, permet à celle-ci de produire davantage de racines secondaires et donc d’absorber de l’eau et des nutriments en plus grande quantité (Coughlan et al. 2000). À l’échelle globale, Maire et al. (2015) ont montré une relation linéaire entre le pH des sols, variant de 3,5 à 8,5, et le taux de photosynthèse net de centaines d’espèces végétales. Le pH est une variable indirecte dans les propriétés chimiques de la fertilité, en régulant la disponibilité des éléments nutritifs du sol pour les plantes (Truog 1948). La quantité totale des éléments nutritifs est un second aspect de la fertilité chimique des sols (Fyllas et al. 2009; Bernier et al. 1989). Parmi tous les éléments nutritifs des plantes, nous nous attarderons davantage sur le N, le P, le potassium (K), le Mg et le Ca. Chacun de ces éléments est essentiel à au moins une des parties du mécanisme de la photosynthèse ou à la régulation de plusieurs systèmes de la plante. Le N et le Mg sont impliqués dans la composition de la Rubisco (Stec 2012; Wang et al. 2017), le P est essentiel dans l’apport en énergie aux différents cycles métaboliques (Veronica et al. 2017), tandis que le K et le Ca sont des régulateurs importants de l’ouverture stomatique (Breck St-Clair 2004; Jin et al. 2011).
Les études reliant fertilité du sol – nutrition des plantes s’ intéressent soit au couple N – P ou à la totalité des bases échangeables des sols (Fyllas et al. 2009; Han et al. 20 Il). À ma connaissance aucune étude n’intègre l’ensemble de ces éléments en essayant de caractériser le rôle de chacun sur les différents mécanismes de la photosynthèse. D’autres éléments nutritifs, comme l’AI et le Mn, sont toxiques en trop grande concentration dans les feuilles (Thornton et al. 1986, St-Clair 2004). Thornton et al. (1986) ont testé l’effet de l’AI à pH faible (4.0) sur la croissance des érables à sucre. Ils ont observé une diminution du contenu en Ca dans les feuilles à mesure que la concentration en AI augmentait. De plus, au-delà de 600 ~M AI, la biomasse des semis diminuait exponentiellement. En Pennsylvanie, la santé des érables et la photosynthèse a été corrélées négativement avec la concentration en Mn des feuilles. L’excès de Mn peut interférer avec l’absorption d’ autres nutriments, entre autres le Mg et le K (St-Clair, 2004). La pratique des amendements cationiques permet à la fois d’augmenter le pH du sol, d’augmenter la totalité des bases échangeables du sol et de limiter ainsi la toxicité en AI et en Mn. Elle serait donc une solution parfaite à la fois au dépérissement des érables et permettrait d’augmenter la croissance de l’érable à sucre. Cette solution a un effet pendant de nombreuses années. Moore et al. (2012) ont observé que 18 ans après l’amendement l’effet des traitements à fortes doses (plus de 2 tlha) était toujours présent sur la concentration en éléments nutritifs (Mg, Ca et Mn) des feuilles, cependant les traitements plus faibles (moins de 2 tlha) n’ influençaient plus l’ absorption.
Propriétés physiques
La structure, la texture, l’humidité du sol, la pente et la profondeur de sol sont des éléments qui peuvent faire varier la fertilité physique du sol. Une bonne profondeur de sol va permettre aux érables de croître et de bien s’enraciner dans le sol afin de résister aux intempéries, ainsi que d’avoir accès à une meilleure réserve d’eau et de nutriments. À plus de 100 cm, on considère que le sol est favorable à la croissance des érables (Brown et al. 2015). Ensuite, la structure du sol joue un rôle essentiel pour conserver l’eau et les minéraux. Un sol qui contient des agrégats, qui sont friables et de faible densité, sera plus propice pour les érables. L’ajout d’amendement vient aider la floculation des éléments ce qui permet de créer davantage d’agrégats (Zhang et al. 2017). Un autre point important dans les propriétés physiques est l’humidité du sol. Le manque d’eau ne semble pas être un problème pour le moment pour nos érablières. Cependant, dans un futur à moyen terme, il est attendu, qu’à cause des changements climatiques, les précipitations, sur les territoires présentement favorables à l’érable à sucre, deviendront de plus en plus erratiques ce qui pourrait rendre son territoire de répartition, en grande partie, défavorable, advenant une diminution de l’ humidité du sol. Ce pronostic a été fait pour les forêts au nord du Minnesota qui sont sur une latitude près de nos forêts (Dymond et al. 2017).
Bien qu’une structure «fertile» permette au sol d’avoir une bonne rétention en eau, il ne faut pas que le sol soit saturé en permanence. Sinon les racines vont être prises dans un milieu anoxique, ce qui limitera le métabolisme racmalre. Pour terminer, la topographie joue un grand rôle dans l’apport et le maintien des nutriments dans le sol. Si la pente est forte, les arbres auront de la difficulté de s’y implanter, le drainage y sera fort, ce qui gardera un faible apport en eau dans le sol et une lixiviation des nutriments. Pour les érables, la pente optimale varie entre 1 et 20 % de dénivelé pour être favorable (Brown et al. 2015). Plusieurs études ont été menées observant toutes que les érables à sucre s’ établissent mieux sur des terrains avec de faibles pentes (St-Clair et q,l. 2005; Arii et al. 2005 et Frey et al. 2007).
Flux hydrique
Le flux hydrique permet aux végétaux de faire parvenir l’eau et les nutriments aux parties aériennes (Figure 1.3 – Partie 3). Le bon fonctionnement de ce flux est essentiel à la survie de n’importe queJ1e espèce végétale. Le stress hydrique influence grandement les plantes, en diminuant la croissance, la teneur en nutriments et la photosynthèse (Naeem et al. 2018). L’un des problèmes rencontrés chez les plantes est le juste équilibre entre la grosseur des vaisseaux et le risque de cavitation. Plus le vaisseau est long et gros, plus il risque d’y avoir la formation d’une buJ1e d’air qui causerait la rupture du flux hydrique et diminuerait l’apport en eau et en nutriment aux parties aériennes de l’arbre. L’une des méthodes afin d’éviter la cavitation est de diminuer la transpiration et donc de diminuer la gs (Sobrado 2009). Il a été suggéré que la gs peut être influencée par quelques variables environnementales. Dans un environnement plus aride, la gs va être plus faible (Farquhar et al. 2002). De plus, il a été démontré en laboratoire et en nature que les plantes limitaient leur conductance stomatique et perdaient moins d’eau avec une plus grande fertilité du sol en azote et en phosphore (Niinemets et al. 2009).
Des études ont réussi à démontrer que l’ajout de Ca sur les végétaux en stress hydrique permet à ceJ1e-ci de mieux survivre aux sécheresses (Li Z. et al. 2017 et Naeem et al. 2018). Cependant, Green et al. (2013) ont testé l’ajout de Ca sur une érablière à bouleau jaune et ont vu une augmentation de l’évapotranspiration de 25 % à 18 % pour les 3 années qui ont suivi l’épandage, démontrant ainsi que l’ajout de Ca pouvait favoriser la transpiration des arbres. Ces résultats semblent donc contradictoires, mais il ressort de là que le calcium aurait une influence importante sur la conductance stomatique. Je pense que le chaulage amène le Ca nécessaire aux érables pour arriver à gérer le manque d’eau dans les mauvaises saisons, ce qui permettrait d’expliquer, en partie, la difficulté de survie des érables à sucre.
Rubisco et son activation
La Ribulose 1,5 – bisphosphate carboxylase/oxygenase, alias la Rubisco, est l’enzyme la plus importante dans le cycle de Calvin (Figure 1.3 – Partie 4). Elle permet de fixer le carbone pour former le sucre que l’arbre utilisera pour ses dépenses énergétiques. Les végétaux ont besoin des cations de Mg pour fabriquer la Rubisco dont le coeur catalytique est supporté par un atome de Mg. De plus, la Rubisco est constituée de 70 % de N. Sans ces deux nutriments, la Rubisco ne pourra pas se fabriquer ou s’activer et ainsi l’arbre ne pourra pas fabriquer de sucre (Stec 2012). Selon les espèces, il est possible d’observer différentes réponses face au traitement de fertilisation. Par exemple, le soja, grâce à des bactéries, augmente son VCrnax avec l’augmentation de la fixation du N dans le sol. Le phénomène est expliqué par une boucle positive entre l’ utilisation de la triose-phosphate et l’activation de la Rubisco (Kaschuk et al. 2012). Alors que pour le saule la fertilisation ne semble pas avoir changé le VCrnax. Dans son cas, la fertilisation a eu un effet sur la surface foliaire qui a fortement augmenté. Pour cette espèce, la stratégie qui a été choisie est d’augmenter ses capacités photosynthétiques par la surface.
Elle a opté pour augmenter la surface en maintenant les mêmes concentrations en Rubisco par centimètre carré (Merilo et al. 2006). Lorsque l’on observe l’effet de la fertilisation sur les érables à sucre, il semble que l’apport en amendements cationiques lui permet d’avoir accès et d’absorber davantage de cations de Mg et de Ca. Nous pensons que l’augmentation de Mg dans les feuilles (Moore et Ouimet 2006), permet à l’arbre de fabriquer davantage de Rubisco et ainsi lui permettrait de produire plus de sucre et ainsi optimiser sa croissance. Dès 1982, Farquhar & Von Cammerer proposent un formalisme mathématique de type Michaelis-Menten pour décrire la relation entre le taux de photosynthèse et la concentration foliaire en Mg. En se basant sur des constantes éparses de la littérature, ces auteurs estimaient que cette relation n’avait qu’une existence physiologique et ont invalidé ce formalisme pour des conditions réelles. In natura, la concentration foliaire en Mg serait toujours suffisante pour activer la Rubisco. Pourtant, cela n’a jamais été testé et semble être une piste suffisamment universelle pour expliquer l’effet résiduel du pH du sol sur la photosynthèse observée à des échelles biogéographiques. À ma connaissance cela n’ajamais été démontré chez l’érable à sucre
REMERCIEMENTS |